

 英語
英語 フランス
フランス スペイン
スペイン ロシア
ロシア 韓国
韓国 日本
日本高分子量ヒアルロン酸粉末とは何ですか?
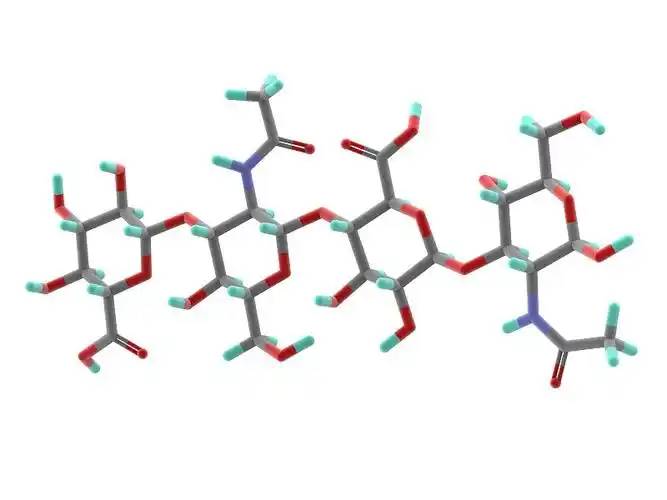
1ヒアルロン酸と構造組成
Hyaluronic酸(HA) is a non-sulfated, high molecular weight linear polysaccharide with a basic structure consisting のa disaccharide unit of D-glucuronic acid とN-acetylglucosamine, linked by a β- 1,3- ligand bond between のD-glucuronic acid and the N-acetylglucosamine, and by a β- 1,4- ligand bond between the disaccharide units (Figure 1). The molecular weight of natural ヒアルロンacid contained でmost tissues ranges from 1000 to 10 000 kDa, and the definition of hyaluronic acid size has not yetbeen fully agreed upon.According to Naro [1], high molecular weight hyaluronic acid is defined as consisting of more than 500 basic 構造units; intermediate molecular weights are generally in the range of 200-500 kDa, and small molecular weight hyaluronic acid is generally defined as less than 200 kDa. According to Monslow etal [2], high molecular weight (HMW hyaluronan) is >1000 kDa, medium molecular weight (MMW hyaluronan) is 250-1000 kDa, low molecular weight (LMW hyaluronan) is >10-250 kDa, and oligo hyaluronan (oligo hyaluronan) is <10 kDa.
The role of hyaluronan in cell motility, cell adhesion and proliferation is mainly mediated by two hyaluronan receptors, CD44 and R-hyaluronan MM (the receptor for hyaluronan-mediated motility, also called CD168). Unlike other mucopolysaccharides, which are synthesised in the Golgi apparatus, hyaluronan is synthesised in the inner membrane of the plasma membrane of the cell by transmembrane synthetases (hyaluronan S1, hyaluronan S2 and hyaluronan S3). Hyaluronan S1 and S2 are responsible for the synthesis of large molecules of hyaluronan and hyaluronan S3 is responsible for the synthesis of small molecules of hyaluronan (<300 kDa), but hyaluronan S3 is more active than hyaluronan S1 and hyaluronan S2 The synthesised large molecules of hyaluronan are cleaved にsmall molecular weights of hyaluronan by ROS (reactive oxygen radicals) and hyaluronan hydrolytic enzyme, and further hydrolysed into low molecular weights of hyaluronan. Further hydrolysis results in oligo- or oligo-hyaluronic acid. Different molecular weights have different physiological functions, e.g. medium molecular weight hyaluronic acid stimulates cell proliferation, while small molecular weight hyaluronic acid promotes cell migration. See Table 1 [3- 19].
2ヒアルロン酸の生物学的進歩
2.1ヒアルロン酸と体重減少
Hyaluronic acid has been approved as a new resource food in China (Ministry of Health Announcement No. 12, 2008), and is used as a food additive and food with health benefits in Korea, as a food additive in Japan, and as a supplement in the United States, Canada, Italy, and Belgium.
Breast milk contains the highest level of hyaluronic acid in the first week of life, which decreases to a stable level over the next two months. In general, a 70 kg adult has about 15 g of hyaluronic acid in his or her body, of which nearly one-third is degraded and synthesised on a daily basis. Hyaluronic acid is taken orally to the 肌and does not accumulate excessively in the body; more than 90% of hyaluronic acid is excreted in the breath or urine [19]. Hyaluronic acid is used for weight loss. When hyaluronic acid was administered to C57BL/6 mice at a dosage of 200 mg/kg for 8 weeks, it was found that hyaluronic acid had a good effect on weight loss, which not only reduced body mass, but also reduced adipose tissue and serum LDL-cholesterol, as well as total cholesterol and leptin, and also reduced adipose tissue hyperplasia, and ameliorated fatty liver. It is therefore hypothesised that the main mechanism of hyaluronic acid weight loss may be through the increase of the peroxisome proliferator-activated receptor PPAR-α and the inhibition of PPAR-γ [20].
2.2ヒアルロン酸と老化
Hyaluronic acid and hyaluronan-binding proteins are involved in cellular senescence. Researchers have found that hyaluronic acid can heal skin damage caused by UV-B exposure through histological changes and wrinkle indicators in vivo. Moreover, high concentrations of hyaluronic acid significantly affected the expression of collagen, matrix metalloproteinase (MMP-1), interleukin IL-1β and interleukin IL-6 protein factors, but not hyaluronan S-2 and transforming growth factor (TGF-β1). Reduction of proteoglycans leads to an increase in free hyaluronan fragments interacting with CD44 protein and phosphorylation of the extracellular signal-regulated kinase ERK1/2, resulting in premature embryonic fibroblast failure [21].

Inhibition of hyaluronan synthesis by 4-methylumbelliferone or targeting of hyaluronan S-2 by miRNA induces cellular senescence, suggesting that hyaluronan plays a role in skin ageing [22]. また、 hyaluronan concentration is greatly reduced in senescent MSCs and in the cell cycle matrix, mainly due to reduced expression of hyaluronidase [23]. Considering the ability of hyaluronan to inhibit fibroblast senescence induced by oxidative stress in vitro [24]. Thus, hyaluronic acid may be involved in ageing during normal and pathological processes. Further investigation of the role of hyaluronic acid in aging in disease states may shed light on the pathogenesis and aid in the development of therapeutic approaches for these diseases.
2.3ヒアルロン酸と成長と発達
Hyaluronan participates in and promotes cell growth through a variety of signalling pathways. In mice in which the hyaluronan gene hyaluronan s2 was knocked out, the heart and blood vessels developed abnormally, and at 9.5 weeks of development, the lack of an intact endocardial cushion resulted in embryonic lethality (TD Camenisch, 2000). Intraperitoneal injections of hyaluronic acid for 3-8 weeks increased the length of villi and depth of the glandular fossa in the small intestine, the depth of the glandular fossa in the colon, and the proliferation of epithelial cells in both. The opposite results were obtained with PEP-1, a short peptide that prevents the binding of hyaluronan to the receptor. These results suggest that endogenous hyaluronan can regulate normal small bowel and colon growth.
Hyaluronic acid and its binding proteins have been shown to play a role in fibroblast proliferation. Hyaluronic acid promotes TGF-β1-mediated fibroblast proliferation, and LMW hyaluronic acid or hyaluronan オリゴ糖などstimulate fibroblast proliferation under a variety of conditions, suggesting that hyaluronic acid plays an important role in tissue fibrosis. Hyaluronic acid fragments induced myofibroblast differentiation, endothelial cell differentiation, and chondrogenic differentiation [25], whereas hyaluronic acid and mouse tumour necrosis factor α-inducible protein 6 inhibited cardiomyocyte differentiation and osteoclast differentiation of 人間bone marrow mesenchymal stem cells, respectively. These data suggest that hyaluronan and hyaluronan-binding proteins regulate cell differentiation in a complex manner and that this differentiation may be related to cell type and microenvironment.
2.4ヒアルロン酸と免疫炎症
Hyaluronic acid and hyaluronan-binding proteins regulate inflammation and the repair of tissue damage by modulating the infiltration of inflammatory cells, the release of inflammatory cytokines and cell migration. Therefore, hyaluronic acid may act as an immunomodulator in human diseases [26]. Synthetic hyaluronic acid has been found to promote the expression of antimicrobial peptides in intestinal epithelial cells. Oral administration of hyaluronic acid from breast milk induced an increase in mouse defensin HβD2 homologue and MuβD3 via Toll-like receptor 4 (TLR4) and CD44 protein, and the addition of hyaluronic acid cytokines to cultured cells showed that colonic mucosal epithelial cells were more resistant to the enteropathogenic bacterium Salmonella typhimurium. Hyaluronic acid regulates cell proliferation and inflammation.
炎症の细胞みずみずしい素肌を活性化させ、金日成interleukin生産・- 6…IL-1腫瘍壊死因子(TNF -α)文書公表を通じてcytokinesと,LMW hyaluronan断片をそれによってhyaluronan合成部分のHB-EGF動作を通じてhyaluronanフィードバックの炎症応答線維芽細胞を含む遷移及び拡散を有する。ヒアルロン酸はrmaらによって報告されている。s.ヒアルロン酸rmaらは、ヒアルロン酸結合クルクミンがケラチノサイトの増殖を促進し、過酸化水素による酸化損傷を減少させ、傷の部位の細胞移動を改善したと報告した[25]。
Hyaluronic acid can be utilised by probiotics. Probiotics can use N-acetyl-D glucosamine, a precursor of hyaluronic acid, as a nutrient, and Kim etal [27] found that Lactobacillus plantarum K8 lysate significantly increased hyaluronic acid secretion from cells. Lactobacillus lysates inhibited the Th2 response induced by interleukin 4 (IL-4)-driven stimulation and increased the Th1 response, and lactobacilli controlling the Th1/Th2 balance contributed to hyaluronic acid induction and the reduction of atopic dermatitis lesions.
Hyaluronic acid and CD44 interact with each other and have the ability to modulate T-cell activation, Th1 production, B-cell activation and regulatory T-cell functions. Recent studies have shown that CD44 functions in the adaptation of regulatory T cells by interacting with galactose agglutinin 9 [28].
Hyaluronic acid and CD44 regulate neutrophil adhesion and recruitment. It is endothelial CD44, not neutrophil CD44, that mediates neutrophil migration, and hyaluronic acid is also capable of promoting the production of inflammatory vesicles in response to injury. In CD44-/- deficient mice, lymphocytes preferentially stayed in lymph nodes and delayed their entry into synovial joints with an inflammatory response compared to wild-type cells. CD44-/- deficient mice die from non-infectious pneumonia injury, a continuous inflammatory response characterised by impaired clearance of apoptotic neutrophils [26]. In addition, hyaluronan fragments can affect dendritic cell maturation, e.g. LWM hyaluronan fragments not only induce dendritic cell maturation but also switch on alloimmunity. In addition, hyaluronan オリゴ糖などare potential activators of dendritic cells, allowing small molecule hyaluronan fragments to promote cell migration and subsequent allergic modifications.
さらに、リポ多糖の天然受容体であるtlr-4は、ヒアルロン酸受容体の一つである。TLR-4発動核転写因子NFタンパク質kB経路を介してこちらになります。1阳诱pro-inflammatory cytokines myeloid-dependent骨髄性分化ファクタMyD88経由で、もう1台はnon-myeloid-dependent増加MyD88経路はinterferon-induced pro-inflammatory遺伝子タイプI interferonsを介して[26]。
2.5ヒアルロン酸と癌
Hyaluronic acid not only acts as a cellular support and hydrophilic matrix, but also regulates cell adhesion, migration, growth and differentiation (TC Laurent, 1992). These properties have led to the involvement of hyaluronic acid in a number of pathological processes. For example, in cancer, hyaluronic acid forms a protective film on the surface of tumour cells, making them less vulnerable to attack by immune cells. At the same time, tumour cells produce more hyaluronic acid or induce the production of hyaluronic acid by releasing growth factors and cytokines. Moreover, ROS-induced hyaluronan fragments contribute to the efficient expression of hyaluronan (R Stern, 2006). Tumour cells and stromal cells are able to express hyaluronan homologues and produce hyaluronan-containing extracellular matrix, which accumulates in the tumour and pericancerous stromal tissues, thus accelerating metastasis of cancer cells [29].
In addition, hyaluronic acid also regulates fibroblast and tumour invasion (Q Yu, 2000). Transforming growth factor and tyrosine kinase, when activated, regulate hyaluronan-mediated invasive cellular motility [27]. Heterogeneity analysis of probe cells using hyaluronan resulted in the identification of subtypes of infiltrating but slow-growing breast tumours. By analysing 线维芽细胞isolated from transgenic mice overexpressing hyaluronan S2, we found that the cells had a greater ability to invade the stroma. If the hyaluronan S2 gene is knocked out of mesenchymal cells, the invasive fibroblast phenotype is terminated, the accumulation of myofibroblasts is discharged and the development of lung fibrosis is inhibited [30].
2.6ヒアルロン酸と幹細胞の相関
The interaction of hyaluronic acid with stem cells has been studied in haematopoietic stem cells, MSCs and pluripotent adult progenitors, suggesting that hyaluronic acid and its binding proteins may interact with various stem cells through their ability to regulate tissue damage and repair processes. During stem cell differentiation, hyaluronan synthesis was enhanced 13-fold and 24-fold, most likely due to an increase in hyaluronan S2 expression. Hyaluronan is required for the production of haematopoietic cells during human embryonic stem cell differentiation. Knockdown of hyaluronan S2 resulted in inhibition of human embryonic stem cell differentiation. Hyaluronan oligosaccharides increase the stem cell properties of epi真皮cells by regulating integrins. Hyaluronic acid promotes CD44-dependent migration of MSCs CD44 has long been used as a marker for stem cells, including embryonic, mesenchymal, haematopoietic and cancer stem cells. In addition, CD44 may play a role in regulating stem cell function. A recent study has shown that although CD44 is not a master stem cell gene, CD44 contributes to stem cell generation and the homeostasis of stem cells in their ecosystem. Hyaluronic acid is required to regulate the haematopoietic support function of bone marrow helper cells and is involved in haematopoietic site assembly.
2.7ヒアルロン酸とアポトーシス
In bleomycin-induced lung injury, hyaluronic acid protects mouse lung epithelial cells from apoptosis. High molecular weight hyaluronic acid reduces UV-induced apoptosis and inflammation in human epithelial corneal cells. Dermal fibroblasts are resistant to stress-induced apoptosis in the presence of high levels of hyaluronan s2 gene expression in the presence of hyaluronan s1/3 knockout, suggesting that hyaluronan S2 protects dermal fibroblasts from environmental stress-induced apoptosis. For inflammatory cells, hyaluronic acid appears to induce apoptosis. Hyaluronic acid appears to induce apoptosis. Hyaluronic acid induces apoptosis in activated T cells via CD44. LMW hyaluronic acid-TLR4 interaction induces apoptosis in inflammatory neutrophils. In rats, administration of hyaluronic acid with a molecular weight of 1600 kDa significantly reduced smoke-induced neutrophil infiltration, pulmonary oedema, respiratory apoptosis and mucus plugging, suggesting that high-molecular-weight hyaluronic acid may have a potential therapeutic effect on smoking-induced lung injury. In addition, cross-linking of CD44 molecule-specific epitopes rapidly induced neutrophil apoptosis in vitro and inhibited neutrophil-dependent renal injury in rats in vivo.
2.8細胞の移動と浸潤における役割
Hyaluronic acid mediates inflammatory cell migration to regulate the inflammatory response and tissue damage.CD44 deficiency leads to increased neutrophil migration and lung injury in E. coli pneumonia in mice. TSG6 is a potent inhibitor of neutrophil migration in an in vivo model of acute inflammation. Exogenous 低分子ヒアルロン酸, or LWM hyaluronic acid produced by overexpression of HYAL1, promotes the migration of dendritic cells from the skin and subsequently alters the allergic response in a TLR4-dependent manner. Hyaluronan and hyaluronan-binding proteins play a role in fibroblast migration and smooth muscle cell migration. Abnormal accumulation of hyaluronan matrix promotes fibroblast migration.

Specifically sized hyaluronan oligosaccharides stimulate fibroblast migration and excisional wound repair. both CD44 and HMMR have been shown to have a role in fibroblast migration during tissue injury. In addition, myofibroblast migration can be regulated by hyaluronic acid. These studies suggest a role for hyaluronic acid in tissue injury and fibrosis. Small molecular weight hyaluronic acid35 and high molecular weight hyaluronic acid117 showed significant differences in the migration and invasion of breast cancer 4T-1 and SKBR3 cells. In healthy people, the level of hyaluronic acid in the body is 10-100 μg/Lbut in breast cancer patients, the level is as high as 200-300 μg/L, and in advanced breast cancer, it reaches 789-2343 μg/L (EH Cooper, 1988). Compared with small molecular weight hyaluronic acid, high molecular weight hyaluronic acid is able to exert a greater squeezing force on the tumour spheres and prevent the growth of cancer cells. Small molecular weight hyaluronic acid can accelerate the process of metastatic fibrosarcoma cell invasion [9].
3展望
Over the past decades, hyaluronic acid and its hyaluronan receptor have been intensively investigated, leading to an increasing use of hyaluronic acid. Nevertheless, there are still some problems that need to be solved. The first problem to be solved is the definition of high molecular weight and low molecular weight hyaluronic acid. There is no international consensus on the distinction between high and low molecular weight hyaluronic acid, which poses a problem in elucidating the physiological effects of hyaluronic acid on cells. Secondly, high molecular weight hyaluronic acid controls the normal homeostasis of the body and shows anti-inflammatory and anti-cancer effects, while low molecular weight hyaluronic acid and oligo-hyaluronic acid show pro-inflammatory and pro-cancer effects. If the same receptor is used, but the results are significantly different, the mechanism is still not clear. In addition, how hyaluronic acid precisely regulates the occurrence and development of inflammation and tumour cells; whether oral administration of hyaluronic acid affects the intestinal flora, which in turn affects human health; and how modified hyaluronic acid (e.g., sulphated hyaluronic acid) has a different mechanism of action compared with non-modified hyaluronic acid, need to be further elucidated. The solution of these problems will certainly help to prevent diseases and maximise the use of hyaluronic acid for the treatment of human diseases.
参照
[ 1] Naor D。 [社説]相互作用 hyaluronic acid and ◆受容体 (cd44、rhamm)は炎症やがんの活性を調節する[j]。正面Immunol、2016」、「7:39。
[2] monslow j, govindaraju p, pure e。Hyaluronan - 機能性や structural 甘い 組織微小環境におけるスポット[j ]。^パウサニアス、2015年6月6日、231頁。
[3] karbownik ms, nowak jz。ヒアルロン酸:新規抗がん治療に向けて[j]。^ a b c d e f g h i(2013年)、105 - 105頁。
[4] papakonstantinou e、roth m、karakiulakis g .ヒアルロン酸:皮膚の老化におけるキー分子[j]。^デルマノ内分泌(2012),4(3):253—258。
[5] kavasi rm, berdiaki a, Spyridaki I, ら皮膚恒常性と炎症性疾患におけるha代謝[j]。^ food chem toxicol, 2017,1 00:1 128- 138。
[6] zhu y, hu j, yu t, etal。高分子ヒアルロン酸は、子宮内膜の線維化を阻害します[j]。医学 ^ a b c d e『仙台市史』、2016年、22 - 34頁。
[7] zhao yf, qiao sp, shi sl, et al。分子量の異なるヒアルロン酸で三次元微小環境を調節すると、乳がん細胞の浸潤挙動が変化する[j]。ACSたら 築城 2017得る。9(11日):9327 - 9338。
[8] noble pw, lake fr, henson pm, et al。ヒアルロン酸によるcd44の活性化は、α依存性の腫瘍壊死因子によるインスリン様成長因子1発現を誘導する 機構 murinemacrophages [J]。^ a b c d e f g h i j j clin invest, 1993,91(6):2368- 2377。
[9]ホッジ- Dufour J 貴族 PW、 ホートン ミスター et アル 誘導 ヒアルロン酸によるil- 12とケモカインの接着依存性プライミングは、居住者ではあるが、誘発されていないマクロファージを必要とする [J]。j immunol, 1997, 159(5): 2492—2500。
[10]大河原泰、田村g、岩崎t, et al。活性化や変形能力- ing増殖因子-ヒアルロン酸による好酸球のβ産生[j]。^ a b c d e f g h i『仙台市史』通史編4(通史編4)444- 451頁。
[11] fitzgerald ka, bowie ag, skeffington bs, et al。ras、プロティンキナーゼc zeta、およびi kappa bキナーゼ1および2は、t- 24癌細胞におけるnf - kappa bbyヒアルロン酸断片の活性化においてcd44の下流のエフェクターである[j]。2000年JImmunol 164(4): 2053 - 2063。
[12] Slevin M クマール S 騒いでる J。 血管の形成 oligosaccharides ヒアルロン酸は血管に影響を与える複数のシグナル経路を誘導する 血管内皮細胞のmitogenicおよび創傷治癒反応 [J]。J Biol ^ a b c d e f g h i(2002) 277(43) 41046- 41059。
[13] west dc, kumar s .ヒアルロン酸と血管新生 [J]。ciba found symp, 1989,143: 187—201;約数の和は187、和は187。
[14]西dc、クマールs .ヒアルロン酸とそのオリゴサカの効果- 内皮細胞の増殖と単層の完全性に乗っています [J]。^ a b c d e f g h i(1989年)、183 - 179頁。
【15位】Vistejnova L, Safrankova B Nesporova K et アル 低分子ヒアルロン酸介在cd44依存的il - 6誘導 感知 in human dermal fibroblasts potentiates 自然免疫応答[j]。^ a b c d e f g h i(2014年)、97- 103頁。
[16] Termeer CC Hennies J Voith U, et アル オリゴ糖など ヒアルロン酸は樹状細胞の強力な活性化因子である [J]。J Immunol 2000年、165(4):明治3年(1870年)3月30日(旧暦)(1863年。
〔17〕Pandey MS。 baggenstoss ba, washburn j, et al。エンドサイトーシスのヒアルロン酸受容体 (hare)は40- 400- kda、しかしそれより小さくても大きくてもないヒアルロン酸に応答して、nf - kappab -介在遺伝子発現を活性化する[j]。^ a b c d e f g h i j biol chem, 2013, 288(20):14068- 14079。
[18] ghatak s, misra s, toole bp。haオリゴ糖は、ホスホイノシチド3キナーゼ/ akt細胞の生存経路を抑制することにより、腫瘍細胞のアンカジ非依存的成長を阻害する [J]。^ a b c d e f g h i j biol chem, 2002, 277(41): 38013- 38020。
[19] oe m, mitsugi k, odanaka w, et al。食事中のヒアルロン酸の移動 into the skin of rats [j]。 科学 世界 日記に、2014年、2014:378024人である。
[20] park bg, park ys, park jw, et al。c57bl /6マウスの高脂肪食におけるenzy-matic ha断片の抗肥満性は肥満を誘発した[j]。^ biochem biophys res commun, 2016,473(1): 290- 295。
[21] suwan k, choocheep k, hatano s, et al。versican / pg- mはヒアルロン酸を細胞外マトリックスに集合させ、胚性線維芽細胞の早期老化に向けたcd44を介したシグナル伝達を阻害する[j]。^ a b c d e f g h i j biol chem, 2009,284(13): 8596- 8604。
。[22]Lompardia SL、 Papademetrio DL Mascaro M et アル ヒト白血病細胞株は、老化を防ぎ化学療法に抵抗するためにヒアルロン酸を合成する[j]。2013年Glycobiology、23(12):1476年(文明8 1463。
[23] jung em, kwon o, kwon ks, et al。相関関係の証拠は-である ヒト間葉系幹細胞の細胞老化期におけるvcam- 1発現の低下とヒアルロン酸合成をtweenにする[j]。biochem biophys res commun, 2011,404(1): 463- 469。
[24]大人Cirillo N Vicidomini A,ジム・メッケイブM, et ヒアルロン酸ベース 複合線維芽細胞を抑える 酸化ストレスによる老化はin vitroで誘発され、in vivoで口腔粘膜炎を防ぐ[j]。j cell physiol, 2015,230(7): 1421- 1429。
[25] sharma m, sahu k, singh sp, et al。外傷治療のためのリハビリ施設 ヒアルロン酸に結合したクミン:in vitroおよびin vivo評価[j]。^「biotechnol」。artif cells nanomed biotechnol, 2017,28:1- 9。
[26]江d,梁j, noble pw。ヒト疾患における免疫調節因子としてのヒアルロン酸[j]。^ a b c d e f g h i(2011)、221- 264頁。
[27] kim h、kim hr、jeong bj、 ら金の経口摂取の効果- chi-由来のラクトバチルス・プランタルムk8は、皮膚に溶解湿らせます- ing。j microb hyaluronic acidiol biotechno [j], 2015,25(1):74- 80。
[28] wu c, thalhamer t, franca rf, et al。ガレクチン- 9- cd44の相互作用は、適応性調節性t細胞の安定性と機能を高める[j]。^ a b c d e f g h i(2014年)247 - 282頁。
[29] Heldin p, basu k, olofsson b, et al。ヒアルロン酸の合成、分解および結合の自由化は乳癌を促進する[j]。j biochem, 2013,154(5): 395- 408。
[30] li y, jiang d, liang j, et al。重度の肺線維症は、インバが必要です- haとcd44によって調節される線維芽細胞の表現型[j]。^ j exp med, 2011, 208(7):1459—1471。




