

 英語
英語 フランス
フランス スペイン
スペイン ロシア
ロシア 韓国
韓国 日本
日本オクタコサノールの研究は、スポーツのパフォーマンスを向上させます
Octacosanol(CH3[CH2]26CH2OH) is a naturally occurring long-chain fatty alcohol that is the main nutrient in the waxy components of plants [1-2]. Recent research has shown that octacosanol is also found in animal food [3]. As a natural nutrient, octacosanol is completely absorbed into the blood and various tissue cells in humans and animals [4] and metabolised by the body through pathways such as fatty acid oxidation [5]. Since the 1990s, the physiological regulatory functions of octacosanol have been increasingly understood and studied. の earliest research found that octacosanol can lower blood lipids [6]. Octacosanol can significantly lower blood cholesterol and low-density lipoprotein (LDL), while increasing high-density lipoprotein (HDL) levels [7].
さらに、オクタコサノールの消費は、血液凝固機能、エネルギー代謝、炎症反応、運動機能、抗酸化能力、神経系機能など、ヒトおよび動物の複数の生理学的プロセスを調節することができ、畜産にも有益な効果があることが現在の研究で明らかにされています。オクタコサノールは低毒性で低コストという利点があるが、その具体的な制御効果とメカニズムについては、さらなる研究が必要である。例えば、オクタコサノールの脂質代謝に対する調節効果が有意でない場合、さらに確認する必要がある。本論文では、オクタコサノールの吸収と代謝、およびその生理的調節作用のメカニズムを概観し、オクタコサノールの研究と応用のための新しいアイデアと方向性を提供する。
1オクタコサノールの源
Octacosanol is the main component of a major natural alcohol mixture isolated from wheat germ oil extract (policosanol), sugar cane extract, rice bran wax and beeswax, and was first extracted from wheat in 1933 [1]. Octacosanol is usually found in the fruits, leaves and seeds of plants, as well as in the waxy components on the surface of plants [8-11]. Studies have shown that the content of octacosanol varies greatly in different plants. For example, the content of octacosanol in perilla oil can reach 400 mg/kg, while only trace amounts are detected in corn oil and soybean oil [2]. Different growth conditions (such as light and growth period) can also affect the content of octacosanol in the same plant [12-13]. High levels of tetracosanol have also been found in animals, for example, Antarctic krill contains about 10.6 µg/mg tetracosanol [3]. In addition, animal foods such as honey, the Chinese medicinal insect Chinese ground beetle, and the lipid-lowering bee waxin also contain tetracosanol [14-16].

オクタコサノールは様々な動植物に含まれていることが分かっていますが、通常の食事で摂取できるオクタコサノールの量は非常に少ないです。したがって、オクタコサノールは通常、その生理学的調節効果を調べるために動物およびヒトの研究で補足されている。
2オクタコサノールの吸収と代謝
コーティングしoctacosanol人や動物に吸収されます。50 mgのオクタコサノールをヒトに投与すると、血液中から無傷のオクタコサノール分子が検出され、完全に循環系に吸収されることが示されています[4]。ラットに14 c標識オクタコサノールを経口投与したところ、血液だけでなく肝臓、脂肪、筋肉、脾臓、腎臓、心臓などの組織からも相当量の放射性物質が検出された。1時間以内に最も高い血中濃度に達し、14 cで標識したオクタコサノールを1回経口投与しても、組織中の放射性物質は最大3日間持続した。オクタコサノールは異なる組織に吸収されるが、吸収量は大きく異なり、脂肪組織(特に褐色脂肪組織)、肝臓、筋肉はより多く吸収する[17-18]。
オクタコサノールの全体的な吸収効率は高くない。オクタコサノールは水に不溶で、室温では食用油にわずかに溶けます。この弱い溶解度はオクタコサノールの吸収効率を低下させる[19-20]。ラットに60 mg/kgのオクタコサノールを投与した研究では、最大血中濃度は30.4 ng/ ml、肝臓では68.4 ng/gであった。アカゲザルに10 mg/kgの経口投与を行ったところ、最大血中濃度は78.2 ng/ mlであった[21]。14 cで標識した2-エチルヘキサノールをラットに経口投与したところ、摂取した2-エチルヘキサノールは投与量の約32%を占め、大量に糞中に排泄された。また、代謝によって生成されたco2と尿から放射能が検出され、尿中の放射能は脂質抽出物ではなく水相抽出物に集中していた[18]ことから、吸収されたオクタコサノールは組織細胞に吸収され、代謝される可能性があることが示唆された。

これを大きくまとめれば、octacosanol[4]の血液を完全には吸収同化されことができやでも様々に吸収ことができる組織細胞octacosanoic酸[21]に改宗することができるまた入室する脂肪酸β-oxidation代謝経路さらに代謝[18 21]。研究によれば、後octacosanolサルのアカゲザルなどの薬を口にある媒体の血中浓度-などの饱和脂肪酸long-chainを与えますオクタデカン酸(C18)はパルミチン酸(C16)とmyristic酸(C14)増え的かつ継続的内容など不飽和脂肪酸オレイン酸(C18)も[21]増えた。このことからoctacosanolを代謝されさせることができるβ-oxidationに短いチェーン脂肪酸に最終的に体なったCO2にエネルギーを提供。これはオクタコサノールが生理的調節効果を発揮する重要な機構であると考えられる。
3オクタコサノールの生理学的調節効果
3.1血中脂質の低下
The first biological function of octacosanol to be discovered was to lower blood lipids. In 1994 [22] and 1995 [6], it was found that supplementing animal food with octacosanol could lower blood lipids such as cholesterol, LDL and triglycerides. Subsequently, a number of human clinical trials also showed that octacosanol can lower blood cholesterol and LDL while raising HDL [7]. Compared to statin drugs, octacosanol has fewer side effects and is safer. It also has a lipid-lowering effect in people who are not already suffering from high blood lipids, so food supplementation with octacosanol can be used as a preventive measure. However, these clinical trials had small sample sizes and short durations, and larger, longer-term clinical trials are needed to truly confirm the lipid-lowering effect of octacosanol.
Early rat experiments showed that octacosanol may inhibit the activity of enzymes involved in triglyceride synthesis [6]. Octacosanol was shown to inhibit cholesterol synthesis by inhibiting the expression of the cholesterol synthesis rate-limiting enzyme 3-hydroxy-3-methylglutaryl coenzyme A (3-Hydroxy-3-Methylglutary Coenzyme A, HMG-CoA) reductase, thereby reducing cholesterol levels [23]. In vitro cell test results show that octacosanol does not directly inhibit the activity of HMG-CoA reductase, but rather exerts a regulatory effect on HMG-CoA reductase by activating AMPK [24-25]. In addition, octacosanol can inhibit the oxidation of LDL, regulate the absorption and clearance of LDL, and thus regulate the level of LDL in the blood [26].
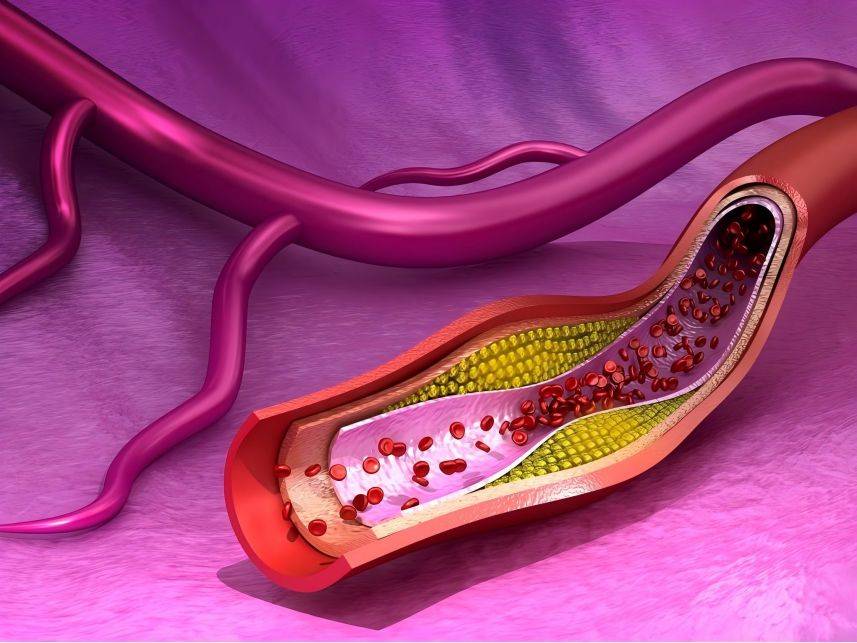
しかし、一部の検査結果によると、オクタコサノールには抗酸化作用はなく[27]、むしろ胆汁酸の吸収を阻害することによってコレステロール値を低下させることが示されている。いくつかの研究では、オクタコサノールは血中脂質を有意に減少させず[28-30]、血中コレステロールおよびldlレベルに影響を与えないが、コレステロール代謝物の排泄を減少させ、全身コレステロール蓄積を減少させることができることが示されている[4]。オクタコサノールの脂質代謝調節における役割については、現在も議論が続いている。サンプルサイズおよび今後の研究の期間を増加させることに加えて、注意もオクタコサノールの吸収およびオクタコサノールの投与量を監視することに注意すべきである。
3.2の血液の凝固を防ぐ
高用量のドコサノール(50 - 200 mg/kg)は、ラットの血小板凝集を阻害し、抗凝固作用を有する[31]。ドコサノールは、血小板から放出される凝固因子であるトロンボキサンa2 (txa2)の形成を阻害することにより、抗凝固作用を発揮する可能性がある[32 - 33]。高脂血症の患者は、一般的に心血管と脳血管の血栓症を起こしやすい。脂質を下げる薬と凝固を同時に調節する薬の開発は、高脂血症などの代謝性疾患を治療するための目標だった。しかし、オクタコサノールの抗凝固作用に関する研究はほとんど行われておらず、その効果とメカニズムについてはさらなる研究と確認が必要である。
3.3エネルギー代謝の調節
いくつかの研究では、オクタコサノールはマウスの高脂肪食品による肥満を抑制することが示されている。高脂肪食では、オクタコサノールを補充することで、マウスの褐色脂肪の遊離脂肪酸受容体4 (ffar4)の発現を増加させ、脂肪組織におけるアンカプリングタンパク1 (ucp1)を介した非震え熱産生を促進し、体内のエネルギー消費を増加させ、体脂肪を減少させることができる[5]。現在、のoctacosanolご入館できません知られるβ-oxidation代謝体にエネルギー供給への通路施設を摂取エネルギー増えている。しかし、それ以上の結果は、オクタコサノールが肥満に対して有意な抑制効果を持つことを示していない。エネルギー代謝の調節におけるオクタコサノールの役割は、さらなる研究が必要である。
3.4消炎効果
炎症応答におけるオクタコサノールの制御的役割も多くの研究分野である。カルダモンの葉は、オクタコサノールや他の脂肪族化合物などの有効成分を含み、伝統的なインドの抗炎症性ハーブです[34]。一生、肉の脂肪Long-chain octacosanolなどから抽出されたオリーブオイル残材[35]やタデmultiflorum葉[36]表情を抑えることができinducible一酸化窒素Synthetase (iNOS)、炎症要素マクロファージリポ多糖類に刺激され(のLPS)刺激マクロファージRAW264.7 inducibleの表現一酸化窒素Synthetase (iNOS)、一酸化窒素の生成を低減しそのことから、自然脂肪一生消炎効果用があります。
アカネ科の植物の葉から抽出したOctacosanolだけでも、白血球数を大幅に減らせる球腫瘍壊死要因のコンテンツを-α(TNF -α)胸膜誘発型宫腔マウスの病気か何かカラギーナンさらにの消炎効果を誇示しOctacosanol(37)。Octacosanolによる大腸炎誘発效果も効率よく解消できデキストラン硫酸ナトリウム(後悔)ラットに濃度を大幅に増大を抑えることが炎症TNF -αなどinterleukin-1β(IL-1β)、インターロイキン- 6誘発型(IL-6)とiNOS後悔肠管組織ましょう。[38]ふたを開けて実験結果追加とoctacosanolをTNF -α颜に調節することができ直接IL-1β、IL-6とiNOSマクロファージ生264.7 MAPK / NF規制され-κB /シグナリング経路AP-1、消炎効果を産地である。
3.5モーター機能の強化
テトラコサノールは運動機能を改善することができる。早くも1960年代には、動物実験により、テトラコサノールの経口投与に抗疲労効果があり、運動能力の効果を高めることが示されました[39-40]。テトラコサノールを補充すると、ラットの連続走行時間が有意に増加する[40]。研究によってoctacosanolの運動くする効果発見された状態では体は大腿特性をoctacosanol補完のほうが改善するでネズミが増加する逆停止や胸腺示唆octacosanol栄养补助食品を用いることができる宇宙飛行士がの損傷を改善body's motor function caused by weightlessness [41]. Studies have found that supplementing with a mixture of octacosanol, branched-chain amino acids, restricted amino acids, carnitine and vitamins can accelerate the recovery of the heart rate of drug-detecting dogs (German shepherds) after exercise and reduce muscle damage caused by exercise [42].
オクタコサノールは運動時間を大幅に増加させることができる。肉体的疲労状態での運動後に補充した場合、オクタコサノール群のラットの血糖および筋肉内グリコーゲン含有量は対照群と有意に変わらなかった[43]。このことは、オクタコサノール補充が筋グリコーゲンの使用を節約することによって運動性能を改善する可能性があることを示唆している。同時に、血中クレアチンホスホキナーゼと筋クエン酸合成酵素の活性は対照群よりも高く、オクタコサノールは筋肉の酸化的リン酸化能力を改善することが示唆された。同位体追跡実験の結果、運動ラットの筋肉組織におけるオクタコサノールの蓄積は、運動していないラットよりも高いことが示された[44]。エネルギー筋の主体になる運動は供給脂肪酸β-oxidation[45]。Octacosanolご入館でき脂肪酸β-oxidation代謝経路[21]のエネルギー产生とそのことから、Octacosanol運動能力を高め筋肉エネルギー供給増えている。また、短期実験結果選手ショーは補完というoctacosanolの活動を増進させることが出来る超酸化物イオンdismutase (SOD)とグルタチオンのperoxidase (GPx)で血液演习后に、強力なmalondialdehydeを减らすとともに局(MDA)は、示唆octacosanolによる酸化ストレス解消ハードな運動[46]。

3.6抗酸化
The antioxidant effect of octacosanol has been found in liver tissue in addition to muscle tissue. In a rat model of acute carbon tetrachloride-induced liver injury, octacosanol supplementation can alleviate the increase in blood transaminase activity and the increase in myeloperoxidase, xanthine oxidase activity, lipid peroxidation (LPO) levels, as well as the reduced activity of SOD and catabolic enzymes and the decreased glutathione content in the liver [47]. Octacosanol can even reduce liver LPO levels and increase glutathione content in normal rats that have not been treated with carbon tetrachloride [47]. In addition, in a mouse model, octacosanol can relieve the stress caused by oxidative stress, thereby improving the harmful effects of stress on sleep [48].
3.7神経系および神経変性疾患に対するオクタコサノールの効果
オクタコサノールを7日間経口投与すると、人間の脳の反応時間を短縮できることが研究で示されており[49]、オクタコサノールは神経学的機能に有益な効果がある可能性が示唆されている。parkinson &のマウスモデルで#疾患6-ヒドロキシドパミン(6- ohda)によって誘発され、マウスにオクタコサノールを14日間添加すると行動障害が改善した。オクタコサノール治療は、黒質と線条体のチロシンヒドロキシレーザー陽性(th陽性)ニューロンの減少を抑制し、線条体のアポトーシスを減少させる[50]。オクタコサノールは神経成長因子(pro-nerve growth factor, ngf)と神経成長因子(nerve growth factor, ngf)の経路を調節することでこれらの効果を生じる。オクタコサノールはまた、1-メチル-4-フェニル-1,2,3,6-テトラヒドロピリジン(1-メチル-4-フェニル-1,2,3,6テトラヒドロピリジン,mptp)誘導マウスのパーキンソン病に対して緩和効果を有する'sモデルには緩和効果もあり、p38分裂促進プロテインキナーゼ(p38 mapk)およびc-jun n末端キナーゼ(jnk)シグナル経路を調節することによって達成される[51]。
4動物生産におけるオクタコサノール
畜産では、オクタコサノールを補うことでブロイラーの生産性を向上させることができる。飼料に24 mg/kgのオクタコサノールを添加すると、ブロイラーの一日の成長速度が大幅に向上し、鶏の乳房肉の生産量などの生産指標も大幅に改善され、オクタコサノールが家畜の生産効率を向上させる安全な添加剤として使用できることが示唆された[52]。オクタコサノールはまた、鶏の卵の生産と卵の品質を向上させることができます[53]。オクタコサノールは卵胞刺激ホルモン受容体(fshr)、黄体形成ホルモン受容体(lh)、プロラクチン受容体(prl)のmrna発現だけでなく、産卵鶏の卵胞刺激ホルモン(fsh)とエストラジオールの血清レベルを増加させる。また、卵巣組織の重量も有意に増加したことから、オクタコサノールはニワトリの生殖能力を向上させ、それによって卵の生産などの生産能力に影響を与えることが示唆される[54]。豚の研究では、オクタコサノールがトリヨードサイロニン(t3)、成長ホルモン(gh)、グルカゴン(gu)、アドレナリン(ad)の血清レベルを調節することが示されている[55]。しかし、ラットおよびウサギを対象とした研究では、オクタコサノールを含む長鎖脂肪酸アルコール混合物は親および子孫に対して毒性がなく、子孫数などの生殖指標に有意な影響を及ぼさないことが示されている[56-57]。したがって、オクタコサノールは畜産分野での応用の見通しが良いかもしれないが、その特定の役割とメカニズムはまだ広範な研究を必要とする。

5まとめと展望
これを大きくまとめれば、octacosanol is a natural nutrient with multiple effective physiological regulatory effects and high safety, and has potential application value in animal husbandry. However, there have been few purification experiments on octacosanol at home and abroad. In the experiments studying its physiological functions, octacosanol is mostly a natural extract and its composition is generally a mixture. Therefore, in future research and applications, the source of octacosanol needs to be considered, more in-depth exploration of its physiological functions and molecular mechanisms, and exploration of the potential characteristics and functions of octacosanol, so as to promote industrial-scale production. It is also possible to further precisely study the nutritional strategies of mixing octacosanol with other long-chain fatty alcohols, providing theoretical guidance for the prevention and treatment of human diseases and the safe and efficient production of animals.
参考:
[1] pollard a、chibnall ac、piper s h .小麦ワックスからのn-octac-オサノールの分離[j]。^「biochem j, 1933, 27(6): 1889-1893」(英語). biochem . j .(1933) . 2018年3月27日閲覧。
[2] jung d m, lee m j, yoon s h, et al。ガスクロマトグラフィー-タンデム四重極質量分析の商業用植物油中ポリコサノールの分析[j]。j food sci, 2011, 76(6): c891-c899。
[3] gao w, liu d, sus。高性能薄膜クロマトグラフィー南極オキアミ(euphausia superbadana)の1-オクタコサノールの定量のための[j]。j chromatogr sci, 2015, 53(5): 811-815。
[4] keller s、gimmler f、jahreis g .オクタコサノールのヒトへの投与は、糞便中の中性ステロラン胆汁酸濃度を低下させる[j]。^ a b c d e f g h i(2008)、109-115頁。
[5] sharma r, matsuzakat, kaushikm k, et al。オクタコサノールとポリコサノールは、褐色脂肪組織を活性化し、肝臓代謝を改善することで、高脂肪による肥満と代謝障害を予防する[j]。sci rep, 2019, 9(1): 5169。
[6] kato s, karinok, hasegawa s, et al。オクタコサノールは、高脂肪食を与えたラットで脂質代謝に影響を与える[j]。1995年Br JNutr 73(3): 433-441。
[7] taylor jc, rapport l, lockwood g b .ヒトの健康におけるオクタコサノール[j]。^ a b c d e f g h i(2003年)19 -19頁。
[8] ganies a、yadavs s . holoptelea integrifolia (roxb)。planch: a review of its ethnobotany, pharmacology, and phytochemistry [j]。^『官報』第401213号、大正14年。
[9] jamkhande pg, bardes r .駆虫活性およびin silicoパスの評価は、cordiadichotoma (forst .)根エキスの予測に役立った[j]。^ anc sci life, 2014, 34(1): 39-43。
[10] iscav m, seca am, pinto d c, et al。食用塩生植物salicornia ramosissimaの親脂性プロファイル[j]。^『仙台市史』通史編、2014年、165 -336頁。
[11] trabelsi h, renaud j, herchi w, et al。チュニジアの3つのピスタシア・レンティカス集団の果実から得られたトリアシルグリセロールおよび脂肪族アルコール[j]。j sci food agric, 2015, 95(10): 2028- 2032。
[12] harrabi s, ferchichi a, bacheli a, et al。policosanol compos-牛乳アザミ(silybium marianuml .)の抗酸化と抗関節炎活動異なる種子成熟段階での油[j]。脂質健康(吉森)2018年、17(1):82。
[13] muthusamy m, kim jh, kim s h, et al。小麦と大麦の芽に含まれる有益なc−グリコシルフラボンおよびポリコサノール含有量の変化[j]。^ a b c d e f g h i(2016年)、9頁。
[14] wang s, ye f .キャピラリーガスクロマトグルによるfenglasuおよびfenglasuカプセル中のtriacontanolandoctactacosanolの決定- aphy[j]。2001年忠遥菜24(10):741-742。
[15] venturelli a, brighenti v, mascolo d, et al。マイクロ波支援技術に基づく、医薬品目的およびそれ以降の蜜蝋ポリコサノールの抽出および精製のための新しい戦略[j]。jpharm biomed anal, 2019, 172: 200-205。
[16] lu y, jiang p . [eupolyphaga sinensis walkerの化学成分][j]。中国中姚座志,1992,17(8):487- 489,512。
[17] kabir y, kimura s .ラットの肝臓および筋肉における(8-14 c)-オクタコサノンの連続投与後の組織分布[j]。^ ann nutr metab, 1995, 39(5): 279-284。
[18] kabir y, kimura s .ラットにおける経口投与オクタコサノールの分布と代謝[j]。1993年アンNutr Metab 37(1): 33-38。
[19] sen g s, ghosh m . octacosanolは修飾されたナノ結晶として物理化学的特性、放出および生物学的利用能を向上させる[j]。eur jpharm biopharm, 2017, 119: 201-214。
[20] chub, quy, huang y, et al。ペグ誘導体オクタコサノールをパクリタキセル送達用ミセル担体として[j]。2016年IntJPharm 500(以上1分け1敗):345-359。
[21] menendez r、marrero d、mas r、et al。オクタコサノール代謝のin vitroおよびin vivo研究[j]。^ a b c d e f『人事興信録』第2版、2005年、113-119頁。
[22] arruzazabala m l, carbajal d, mas r, et al。コレステロール-ウサギにおけるポリコサノールの低下効果[j]。^ a b c d e f g h i l l res, 1994, 27(3-4): 205-208。
[23] menendez r、amor a m、rodeiro i, et al。ポリコサノールは、培養線維芽細胞におけるhmg-coa還元酵素活性を調節する[j]。^ a b c d e f g h i(2001)、32頁。
[24] singh d k、li l、porter t d . policosanolは、ampキナーゼの活性化によって肝細胞におけるコレステロール合成を阻害する[j]。^ a b c d e f g h i j pharmacol exp ther, 2006, 318(3): 1020-1026。
[25] raje, woos y,ニラs, etal。アデノシン、アデノシンとも表記される 5'-一リン酸活性化プロテインキナーゼ(ampk)の栽培品種と成長時期別の活性化ポテンシャル[j]。food chem, 2020, 317: 126388。
[26] menendez r, fraga v, amor am, et al。ポリコサノールの経口投与はin vitroで銅イオン誘導ラットリポタンパク質過酸化を阻害する[j]。Physiol Behav、 1999年、67(1面)するよ
[27] ng c h, leung k y, huang y, et al。ポリコサノールは、ヒト低密度リポタンパク質には抗酸化活性を持たないが、ハムスターの胆汁酸の排泄を増加させる[j]。^ jagric food chem, 2005, 53(16): 6289-6293。
[28] kassis an, marinangeli c p, jain d, et al。ゴールデンシリアハムスターの血漿コレステロールに対するサトウキビポリコサノールの効果の欠如[j]。^アポロドーロス、2007年(1)、153-158頁。
[29] liny, rudrum m, van der wielen r p, et al。小麦胚芽ポリコサノールは、コレステロール濃度が正常から軽度に上昇した被験者で血漿コレステロールを低下させることができなかった[j]。メタ- ^パウサニアス、2004年(平成16年)10月10日:1309-1314。
[30] dullens s p, mensink r p, bragt m c, et al。ヘテロ接合型ldl受容体欠損マウスにおける、異なる鎖長を有する乳化ポリコサノールのコレステロール代謝への影響[j]。j lipid res, 2008, 49(4): 790-796。
[31] arruzazabala ml, carbajal d, mas r, et al。ラットにおける血小板凝集に対するpolicos - anolの影響[j]。^『仙台市史』通史編3(通史3)321-327頁。
[32] arruzazabala ml, molina v, carbajal d, et al。モンゴルのgerbilsにおける脳虚血に対するpolic- osanolの効果:プロスタサイクリンおよびトロンボキサンa2の役割[j]。prostaglandins leukot essent fatty acids, 1993, 49(3): 695-697。
[33] molina v, arruzazabala m l, carbajal d, et al。モンゴルの生殖器における脳虚血に対するポリコサノールの効果[j]。^ a b c d e f g h i『人事興信録』第32版、1269-1276頁。
[34] sharma p, dwivedee b p, bishtd, et al。cordifolia tinosporaの化学成分と多様な薬理学的重要性[j]。heliyon, 2019, 5(9): e2437。
[35] fernandez-arche a, marquez-martina, dela puerta v r, et al。pomaceオリーブオイルからの長鎖脂肪酸アルコールは、炎症誘発性メディエーターの放出を調節する[j]。jnutr biochem, 2009, 20(3): 155-162。
[36] rodrigues m j, custodiol, lopes a, et al。ポリゴのin vitro抗炎症性および抗糖尿病性の可能性を明らかにする- num maritimum [J]。2017年Pharm Biol、55(1):1348-1357。
[37]de oliveira am, conserval m, de souza f j, et al。antinocic—sabicea grisea var. griseaの葉からのオクタコサノールのeptiveおよび抗炎症作用[j]。^ a b c d e f g h i j j sci, 2012, 13(2): 1598-1611。
[38] guo t, lin q, li x, et al。オクタコサノールは、raw264.7マクロファージと大腸炎のマウスモデルの両方で炎症を抑制する[j]。jagric food chem, 2017, 65(18): 3647-3658。
[39] consolazio c f, matoushl o, nelson ra, et al。潜在的な抗疲労薬の生理学的および生化学的評価。III .オクタコサノール、小麦胚芽油、ビタミンeが遊泳ラットのパフォーマンスに及ぼす影響[j]。^ ab c d e f g h『日本海軍史』第1巻、新潮社、1963年、11 - 12頁。
[40] consolazio c f, matoushl o, nelson r a, et al。オクタコサノール、小麦胚芽油、ビタミンeの遊泳ラットのパフォーマンスへの影響[j]。^ a b c d e f g h i j applphysiol, 1964, 19: 265-267。
[41] kim h, parks, hand s, et al。オクタコサノール補給は、訓練されたラットにおいて、走行耐久時間を増加させ、疲労後の生化学的パラメータを改善する[j]。j med food, 2003, 6(4): 345-351。
[42] bai s, xiel, liu c, et al。食品中のオクタコサノールの影響尾部浮遊ラットにおける物理パラメータ[j]。space med med eng (beijing), 1997, 10(6): 450-452。
[43] menchetti l, guelfi g, speranza r, et al。薬物検出トレーニングコース中のジャーマンシェパードの体力に対する栄養補助食品の利点[j]。plos one, 2019, 14(6): e218275。
[44] kabir y, kimura s .ラットにおける運動に対する放射性オクタコサノールの分布[j]。^『仙台市史』通史編4(通史)373-377頁。
[45] gowans g j, hawley sa, ross fa, et al。ampisは、アロステリック活性化とネットリン酸化の両方を促進することにより、amp活性化プロテインキナーゼの真のphy遺伝子制御因子である[j]。セル ^『仙台市史』通史編4(通史編5)556-566頁。
[46] lee s h, scotts d, pekas e j, et al。テコンドー訓練後のオクタコサノールによる脂質の改善と酸化ストレスの軽減[j]。intj sports physiol perform, 2019, 14(9): 1-7。
[47] ohta y, ohashi k, matsura t, et al。オクタコサノールは、四塩化炭素中毒ラットにおいて、急性肝障害進行に伴う肝活性酸素種の破壊された代謝を減衰させる[j]。j clin biochem nutr, 2008, 42(2): 118-125。
[48] kaushikm k, aritake k, takeuchi a, et al。オクタコサノールはマウスのストレスを緩和することで、ストレスに影響された睡眠を回復させる[j]。sci rep, 2017, 7(1): 8892。
[49] fontani g, maffei d, lodi l . policosanol,反応時間および事象関連電位[j]。2000年Neuropsychobiology 41(3): 158-165。
[50] wang t, liu yy, wang x, et al。prongfおよびngfシグナル伝達の調節を介したラットにおける6-ヒドロキシドパミン誘発性パーキンソン病に対するオクタコサノールの保護作用[j]。^『仙台市史』通史編7(仙台市)765-774頁。
[51] wang t, liu y, yang n, et al。1-メチル-4-フェニル-1,2,3,6テトラヒドロピリジンにおけるオクタコサノールの抗パーキンソン病効果- ネズミか[J]扱われた。^ a b c d e f g h『官報』第1087号、大正7年(1918年)10月10日。
[52] long l, wu s g, yuan f, et al。ブロイラー雛の成長性能、死骸特性および肉の品質に対する食餌性オクタコサノールの影響[j]。^ a b c d e f g h i j anim sci, 2016, 29(10): 1470-1476。
[53] peng k, long l, wang y, et al。米ぬかから抽出したオクタコサノールが産卵鶏の産卵性能、卵の品質および血液代謝物に及ぼす影響[j]。^ a b c d e f g h i j j anim sci, 2016, 29(10): 1458-1463。
[54] long l, wu s g, yuan f, et al。経口オクタコサノールの影響 産卵性能、卵の品質、血清ホルモンレベル、およびthereprodに関連する遺伝子の発現に関する補足- uctive軸象眼鶏[j]。2017年Poult Sci、96(4):894-903。
[55] long l, wu s, sun j, et al。米ぬかから抽出したオクタコサノールの血中ホルモン濃度および遺伝子への影響 離乳子のグルコーストランスポータータンパク-4およびアデノシン一リン酸タンパク質キナーゼの発現[j]。^『仙台市史』通史編1(通史編2)293-298頁。
[56] rodriguez m d, garcia h .ラットおよびウサギにおけるポリコサノールの奇形形成および生殖研究[j]。teratogcarcinog mutagen, 1994, 14(3): 107-113。
[57] rodriguez m d, sanchez m, garcia h .ラットにおけるポリコサノールの多世代repro- duction study of policosanol in rats[j]。^ a b c d e f g h ic d e f g h i(1997年)、97-106頁。




