

 英語
英語 フランス
フランス スペイン
スペイン ロシア
ロシア 韓国
韓国 日本
日本ヒアルロン酸の研究は、メラニン代謝を調節します
異常なメラニンの合成と代謝は、肝斑やその他の色素性疾患の発生の主なメカニズムであり、光老化皮膚の重要な特徴の一つでもあります[1-2]。人々の経済発展と改善と#39;s living standard, there is an increasing demとためhealthy とyouthful skin, とhow にbetter regulate melanでmetabolism is のgreat clinical significance. ヒアルロン酸 has a wide range のbiomedical applications due to ◆good tolerance とnon-immunogenicity. In recent years, it has been used as a filler でaesthetic medicine, playing an important role でfacial rejuvenatiにとimprovement の肌imperfections[3] . Currently, little research has been reported on the relationship between ヒアルロンacid and melanでmetabolism. Whether hyaluronic acid can regulate melanin metabolism deserves in-depth study. In this paper, we will review the roles のhyaluronic acid and cells involved in melanin biometabolism in recent years, so as to clarify the correlation between hyaluronic acid and melanin biometabolism, and to provide information for further research.
1ヒアルロン酸の生物学的特性
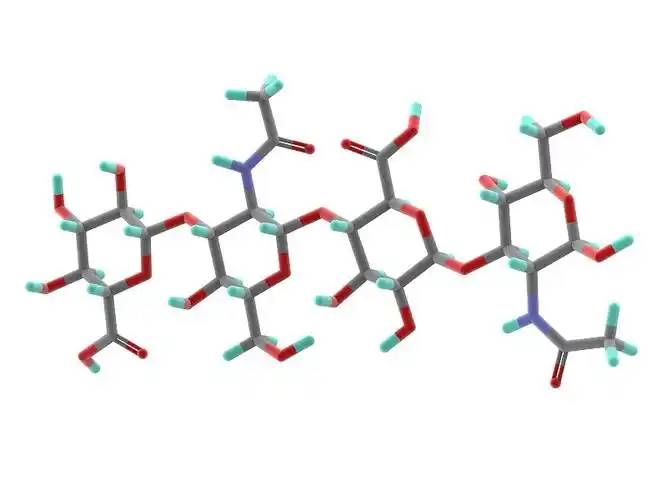
Hyaluronic acid is a non-sulfated glycosaminoglycan composed of repeating disaccharide units of D-glucuronic acid and N-acetylglucosamine. It has a wide range of 分子weights, and its concentration and distribution in the 人間body vary according to tissue type, age, and disease status[4] . Hyaluronic acid acts as a lubricant and shock absorber in the skin and joints due to its viscosity, elasticity and other rheological properties[5] ; it also participates in the regulation of cellular 機能by activating intracellular シグナリングpathways を通じてbinding to hyaluronic acid receptors (e.g., CD44) on the cell surface. The size of the hyaluronic acid molecule is a major determinant of the activation of receptor-mediated signalling pathways, and only hyaluronic acid chains of a specific mass range are able to activate the corresponding receptor-mediated cell signalling, which is regulated by the balance between hyaluronic acid biosynthesis and degradation [4,6].
2メラニンの生体内代謝は様々な要因によって調節されています。
メラニンはメラノサイトで合成され、ケラチノサイトに輸送され、メラノサイトの増殖や生存、樹状突起の形成、メラニン合成、メラノソームの輸送に影響を与えることにより、細胞内および細胞外の様々な因子が、メラニンの生物学的代謝の制御に、プラスまたはマイナスのいずれかに関与しています[7-8]。
炎症と酸化ストレスはメラニン合成に影響を与える:ポスト炎症性色素沈着では、メラニン細胞は、細胞増殖と活性を増加させることによって、炎症に応答し、メラニン産生が増加し、樹状突起を介して近隣のケラチノサイトに転送され、過剰色素沈着をもたらす[9];紫外線は細胞膜の脂質過酸化を引き起こし、メラノサイトの活性酸素種(ros)のレベルを上昇させ、メラノサイトの高色素沈着の生成を刺激する可能性があります[10]。これはメラニン細胞を刺激して大量のメラニンを生成させる可能性があります[10];白斑の発症機序では、rosが分子および小器官の機能不全を引き起こし、さらなる免疫応答を引き起こし、最終的には色素細胞死に至ることが広く受け入れられている[11]。
ケラチノサイト、線維芽細胞、免疫細胞などのメラノサイトに隣接する細胞は、パラクリン因子の分泌を通じてメラニンの代謝調節に関与しています[8,12-13]。色素性皮膚疾患は、しばしば複数の色素細胞隣接細胞由来のサイトカインの異常発現を伴う:黒点の色素沈着領域の表皮では、ケラチノサイト由来の幹細胞因子(scf)およびエンドセリン-1の発現が亢進している[14];肝斑の高色素性皮膚病変では、線維芽細胞由来の分泌性frizzled-associated protein 2 (sfrp2)の発現が有意に上昇している【15位】。(sfrp2)は、線維芽細胞由来の分泌性フリズルド関連タンパク質であり、有意に上昇している[15]。伝統的なparacrine要因としてkeratinocyte-derivedα-MSH、基本線維芽細胞成長因子(FGF-2) fibroblast-derivedケラチン生成細胞成長因子(KGF)や分泌されるタンパク质Dickkopf-1 (DKK1)等が[13、16 ~ 18]ある。ヒスタミン、プロスタグランジンe2、il-6、il-17、腫瘍壊死因子(tnf)などの様々な炎症性メディエーターもメラニン生成の制御に関与している[19-20]。近年,メラノサイト近傍の細胞の傍分泌機能に関与する因子がますます多くなっていることが判明しています[21-23]。
3 .ヒアルロン酸がメラニン生物学的代謝を制御する可能性のある機構
Lim et al[24] designed a self-controlled study using hyaluronic acid to fill the tear troughs. 4 weeks after topical 注射of hyaluronic acid, a decrease in localised melanin content as measured by a skin melanometer was observed compared to the control side, suggesting the potential of hyaluronic acid to influence melanin biometabolism.
3.1メラニン細胞に対するヒアルロン酸の直接の影響
それが判明しました0.2-5 g/ lヒアルロン酸メラニン細胞の増殖を促進して、メラニン細胞のチロシナーゼ活性を増加させて、メラニンの合成を促進することができます;10 g/ lヒアルロン酸はメラニン細胞の増殖を阻害し、チロシナーゼの活性を低下させ、メラニンの合成を阻害します[25]。
Hyaluronic acid has significant anti-inflammatory and antioxidant effects[26-27] : some studies have suggested that低分子ヒアルロン酸induces inflammation, while high molecular weight hyaluronic acid has an anti-inflammatory effect.26 Takabe et al.[28] found that UV treatment of melanocytes resulted in the inhibition of hyaluronic acid synthesis and the expression of the receptor CD44, and the inhibition of IL6, IL8, CXCL1, CXL1, and CXL1 感覚器官melanocytes, IL6, IL8, CXCL1 とCXCL10release in melanocytes; UV light and hyaluronanhydrolase co-treatment of melanocytes increased the release of the above inflammatory factors; exogenous addition of hyaluronic acid tablets did not significantly alter the 生産of inflammatory factors; the above results suggested that the unhydrolysed hyaluronic acid, which was present in the periphery of the melanocytes, could play a protective role in the inflammatory response of melanocytes, but the study did not focus on the role of the hydrolysis in this process in the melanocytic inflammation. The results suggest that hyaluronic acid, which is present around melanocytes and is not hydrolysed, could play a protective role in the inflammatory response of melanocytes, but the study did not focus on whether melanin synthesis was affected in this process. In the skin, hyaluronic acid acts as an antioxidant and free radical inhibitor[29] .
In our previous study, we observed that hyaluronic acid intervention down-regulated ROS levels in 正常and aged fibroblasts, and Campo et al.[30] found that the addition of hyaluronic acid to the 酸化system induced by Fe2+ and ascorbate in fibroblasts inhibited lipid peroxidation through chelation, suppressed ROS production, reduced DNA fragmentation and protein oxidation, and limited cellular growth through the addition of hyaluronic acid. The addition of hyaluronic acid to the ascorbate-inducedROS-generating oxidative system can inhibit lipid peroxidation through chelation, inhibit ROS production, reduce DNA fragmentation and protein oxidation, and limit cell death, thus exerting antioxidant effects. It is not known whether hyaluronic acid can directly exert an antioxidant effect on melanocytes in the current study, and further investigation is needed.
3.2メラノサイト近傍細胞のパラクリン機能制御におけるヒアルロン酸の関与
In previous studies, hyaluronic acid has attracted much attention because of its role in inflammation and wound healing[26,32] . Recently, it has been found that hyaluronic acid can regulate the expression and secretion of growth factors and interleukins by acting on keratinocytes, fibroblasts and immune cells in the skin.
3.2.1ケラチノサイトのパラクリン作用に対するヒアルロン酸の影響
In the keratinocytes of aging mice, the binding of hyaluronic acid fragments (no more than 27 kDa) to its receptor CD44促進RhoA-ROK signalling, while the interaction between high molecular hyaluronic acid (700 ~ 1 000 kDa) and CD44 を刺激activation of Rac-PKNγ, which affects the synthesis and secretion of keratinocytes [33]. UVB irradiation induces the degradation of hyaluronic acid into smaller fragments and selectively 発動different cellular signalling mediated by CD44 in keratinocytes[34] .
In vitro studies have shown that 1% of high molecular hyaluronic acid (700-1,700 kDa) can increase the secretion of α-MSH from keratinocytes into the supernatant[35] . Hyaluronic acid oligosaccharides inhibited the release of IL-8 and TNF-α from LL-37-treated keratinocytes[36] . Hu et al.[37] found that very low molecular hyaluronic acid (0.8 kDa) and high molecular hyaluronic acid (1,200 kDa) inhibited the secretion of IL-6, IL-8, and IL-1 β by 人間表皮keratinocytes after UVB irradiation, and the process involves the effect of hyaluronic acid on the secretion of TLRS and IL-1β in human epi真皮keratinocytes[38] . This process involves the blockage of TLR4 activation by hyaluronic acid.
3.2.2線維芽細胞のパラクリン作用に対するヒアルロン酸の影響
David-Raoudi et al.[38] investigated the 効果of natural hyaluronic acid, 12-and 880-saccharide hyaluronic acid fragments on the proliferation of human fibroblasts and the expression of matrix-related genes, and found that all three types of hyaluronic acid could promote the adhesion and proliferation of cells, and increase the expression of 行列metalloproteinase-1 and matrix metalloproteinase-3, among which 12-saccharide hyaluronic acid could enhance the expression of type I collagen and TGFF-3 in fibroblasts. In particular, 12-saccharide hyaluronic acid enhances the expression of fibroblast type I collagen and TGF-β1.
Ciccone et al.39 reported that FGF-2 expression by fibroblasts was up-regulated when fibroblasts were exposed to 1 mg/mL of soluble hyaluronic acid, whereas the same concentration of a mixture of reticulated and free low-molecular-weight hyaluronic acid did not result in such an alteration.Asparuhova et al.[40] found that hyaluronic acid at 2,500 kDa (4 mg/mL) significantly increased the expression of human oral fibroblasts. Asparuhova et al.[40] found that 2,500 kDa of hyaluronic acid (4 mg/mL) significantly increased the expression of FGF-2, EGF2, IL-1 α, IL-1 β, and TNF in fibroblasts of human oral origin, and Quan et al.[41] showed an increase in the number of fibroblasts in the dermis, elongation of their morphology, and an increase in CCN2 in the dermis by subcutaneous injection of hyaluronic acid.
Radrezzaet al.[42] applied 量子化プロテオミクスto study the changes in protein profiles of normal human dermal fibroblasts with different concentrations of low molecular weight hyaluronic acid in the range of 20-50 kDa (0.125%, 0.25%, and 0.50%) and found that treatment with low molecular weight hyaluronic acid promoted the growth and proliferation of the cells, and increased the biosynthesis of proteoglycans, and the treatment of low molecular weight hyaluronic acid promoted cell growth and proliferation, and increased protein glycan biosynthesis at the highest concentration (0.50%). At the highest concentration (0.50%), inflammatory and immune responses were activated and the secretion of interleukins (IL-12, IL-1, IL-2, IL-4, etc.) and TNF-α was increased.
Lorén et al.[43] found that a 4.3-kDa hyaluronic acid fragment promoted the release of IL-6 and IL-8 from dermal fibroblasts を介してthe CD44 receptor, but Olsson et al.[44] found that no increase in the release of IL-1 β, IL-6, IL-8, IL-10, IL-12, or TNF-α was detected after hyaluronic acid stimulation of fibroblasts from synovial membranes. IL-1β, IL-6, IL-8, IL-10, IL-12 or TNF-α were not detected after hyaluronic acid stimulation.
3. 2. 3免疫細胞のパラクリン作用に対するヒアルロン酸の効果
免疫細胞が分泌する炎症性因子もメラニン合成の制御に重要な役割を果たしています。発見しヒアルロン酸の分泌を抑制するかもしれないIL-6 TNF -αにLPS-inducedに264.7細胞ヘルパーt細胞に使うと low molecular weight hyaluronic acid 主にIFN -γの分泌を抑制、高分子体重ヒアルロン酸は抑制されるIL-4レベル[45]。
3. 2. 4ヒアルロン酸は隣接細胞のパラクリン作用を調節することで、メラノサイトの機能を調節しています
On the basis of the previous theory, we designed a fibroblast-melanocyteco-culture system, and after photo-aging of fibroblasts from human skin, we found that the intervention of hyaluronic acid at 1 mg/mL on photo-aged fibroblasts inhibited melanin synthesis in melanocytes in the co-culture system, but did not have any significant effect on the melanin synthesis in the normal fibroblast-melanocyte system, which preliminarily proved that under certain conditions, hyaluronic acid inhibits the synthesis of melanin in fibroblasts and melanocytes. Under certain conditions, hyaluronic acid can regulate the 機能of melanocytes by regulating the paracrine action of fibroblasts.
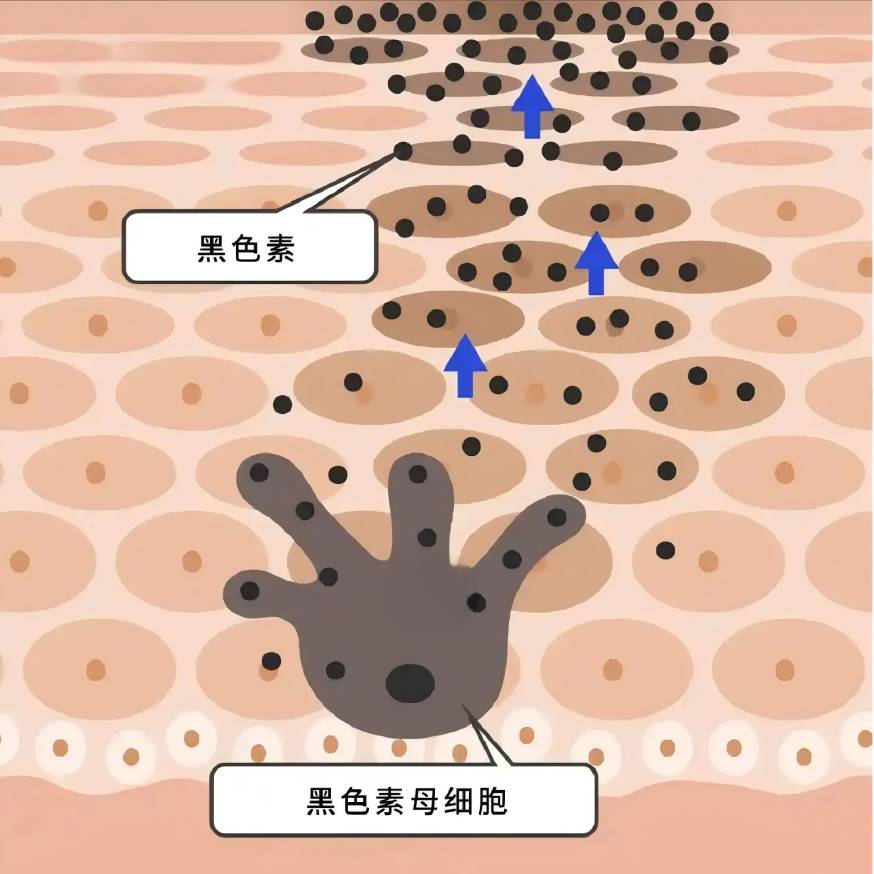
周季常等[46]のUVB照射がIL-18の生産を誘導みずみずしい素肌IFN -投与-かつ時間にγ、メラニンを著しく増加しIL-18コンテンツみずみずしい素肌とco-culturedメラニン色素が多く、IFN -γは著明な抑制効果を示した。D'agostino et al [47] found that a mixture of hyaluronic acid and chondroitin promoted the increase of melanin synthesis in keratinocyte-melanocyte co-cultures, but chondroitin or hyaluronic acid alone did not have a significant effect on melanin content. This may suggest that hyaluronic acid needs to be in a certain form or under a certain condition in order to play its role in regulating melanin synthesis.
4概要と展望
Although hyaluronic acid has been used for a long time and in a wide range of applications, its effects are not well understood because they are affected by a variety of factors such as molecular weight, cross-linking degree, concentration, receptor type, target cell type, etc. In the field of dermatology, hyaluronic acid has been used for a long time and in a wide range of applications. In the field of dermatology, few studies have been reported on the effects of hyaluronic acid on melanin biometabolism. The results of the existing clinical and basic studies suggest that hyaluronic acid may be related to melanin metabolism, especially the metabolic changes of melanin under external stimuli (e.g., UV irradiation), which is worthy of in-depth study. A deeper understanding of the functions of hyaluronic acid適切なヒアルロン酸を選択し、美容医学や色素性疾患の治療においてその機能を実現するのに役立ちます。

参照
[1] Passeron T,接触禁止 M.Melasma、 photoaging 障害[J]。豚今mentて黒色腫Res publica 31(4): 461-465。、2018年
[2] han a, chien al, kang s . photoaging [j]。2014年Dermatol Clin、 32(3): 291-299。
[3] bukhari sna, roswandi nl, waqas m,et バイオメディシン(biomedicine):最近の更新のレビューを有望な皮膚若返り 化粧品およびヌトリコスの前臨床および臨床調査-メティック効果[j]。Int J Biol Macromol、2018年、120、 ^ a b『人事興信録』1682頁 1695年。
[4] KobayashiT、ChanmeeT板野 N.Hyaluronan:谢と机能か[J]。生体物質、2020年(1525 10(11)。
[5]カウマンmk, lee hg, schwertfeger kl,他。コンテンツとサイズ hyaluronanの 生物 流体 and 組織か[J]いない前 Immunol、 2015年6:261ました
[6] weigel ph.ヒアルロン酸のサイズ依存性と特異性を評価するための受容体シグナル伝達研究の計画、評価、検証[j]。glyco - biology,2017,27(9):796-799。
【7】 D'mello sa, finlay gj, baguley bc,et al。^ a b c d e f g h i j j sci,2016,17(7):1144。
[8] pillaiyar t, manickam m, jung sh.recent development of シグナリングpathways inhibitors of melanogenesis[j]メラトニン生成を阻害するシグナル伝達経路の開発[j]。^ a b c d e『官報』第40:9 - 11。
[9] silpa-archa n, kohli i, chaowattanapanit s,et al hyperpigmentation: a comprehensive overview: epidemiology,patho- genesis,clinical presentation,and noninvasive assessment technique [J]。^ a b c d e f『仙台市史』、2017年(平成29年)、591-605頁。
[10] シーズh,シュタールw .栄養保護 反対 skin 傷日光か[J]んだ杏奴牧師さんNutr 24:173-200 04
[11] xuan y, yang y, xiang l,et al.白斑の病因における酸化ストレスの役割:a 犯人 for melanocyte 死か[J]。oxid med cell longev,2022,2022:8498472。
[12] decean h, perde-schrepler m, tatomir c,et al pro-inflammatory cytokines and matrix metalloproteinases produc - tion in co-cultivated ヒトケラチノサイトとメラノサイト[j]。 ^「ararch dermatol res,2013,305(8):705-714」(英語). ararch dermatol res . 2013年5月5日閲覧。
[13]元XH)、ZH。メラニン生成のパラクリン調節[j]。^ a b c d e f『仙台市史』、2018年、178頁(3)。
[14] bastonini e, kovacs d, picardo m .皮膚の色素沈着とピグメン-証人障害:表皮/皮膚のクロストークに焦点を当てる[j]。2016年アンDer - matol 28(3): 279-289。
[15] kim m, han jh, kim jh,et al. frizzled-related protein 2 (sfrp2)メラニン生成刺激剤として機能する;sfrp2の役割 紫外線誘発性高色素性疾患[j]。J Dermatol投資 2016年136(1):236-244。
[16] jiang l, huang j, lu j,et al.霊芝多糖類 il-6 / stat3 / fgf2経路を介して、角化細胞および線維芽細胞のパラクリン作用を阻害することによってメラニン生成を減少させる[j]。Jセル 2019年Physiol 234(12): 22799-22808。
〔17〕 kovacs d, cardinali g, aspite n,et al.線維芽細胞由来の役割 太陽レンズの色素沈着亢進を調節する成長因子[j]。 br j dermatol,2010,163(5):1020 -1027。
[18] ^ a b c d e f g h i、c d e f g h i。本質的・非本質的な規制です ヒト皮膚メラニン生成および色素沈着[j]。国際jコスメsci 40、2018年(4):328-347。
[19] fu c, chen j, lu j,et al (審査)か[J]。Mol 医学 21、2020関心が 1421年- 1430.
[20] yoshida m, takahashi y, inoue s .ヒスタミンはh2を介してプロテインキナーゼaの活性化によってメラノゲン- sisおよび形態変化を引き起こす receptors in human normal メラニン色素が多く[J]。J投資 Dermatol、 2000年114(2):334-342。
[21] yoon je, kim y, kwon s,et al 色素沈着:老人性レンズレンズの治療標的となる可能性[j]。 ^ a b cアポロドーロス、2018年8月8日、17頁。
。[22] 金 Y、康 B、金JC et al.Senescent fibroblast-derived gdf15は皮膚の色素沈着を誘導する[j]。J 投資 Dermatol、2020年、 140(12): 2478-2486。
[23] xu z, chen l, jiang m,et al. ccn1 / cyr61は、ヒト表皮メラノサイトにおいてインテグリンα6beta1、p38 mapk、およびerk1 /2シグナル伝達経路を介してメラノーゲンの生成を刺激する[j]。^ a b c d e f g h i j j invest der - matol,2018年138(8):1825 -1833。
[24] lim hk, suh dh, lee sj,et al injection on nasojugal 溝:予備 無作為 split face clinical controlled study[j]。j cosmet laser ther,2014,16(1):32-36。
[25] hong wei-song, qian guo-pei, xu ai-e。メラノサイトの生物活性に対するガラス酸ナトリウムの影響[j]。メラノサイトの生物活性に対するガラス酸ナトリウムの影響[j]。中国皮膚科学会誌,2011,44(7):491-493。
[26] muto j, sayama k, gallo rl,et al.皮膚生物学におけるヒアルロン酸のes細胞的役割に関する新たな証拠[j]。J索引(Sci) Dermatol 94セットの(1):2019年190 -195。
[27] litwiniuk m, krejner a, speyrer ms,et al。2016年傷は、28 (3):78 - 88。
[28] takabe p, karna r, rauhala l,et al 断片化はuvbによって誘導されるtlr-4受容体のシグナル伝達を増強する と表情 of proinflammatory 者 IL6、IL8 CXCL1、 and CXCL10 via NF-kappaB 活性化[J]。J Dermatol投資し 2019年139人が(9):1993-2003。
[29] マリノa、ヌネスc、レイスs .ヒアルロン酸:主成分 療法 of 炎症か[J]。 生体物質、 2021年まで 15年(1518年)11(10):。
[30] campo gm, avenoso a, campo s,et al.ヒアルロン酸によるdnaフラグの減少とヒドロキシルラジカル産生と chondroitin-4-sulphate in 鉄 プラス ascorbate-induced oxidative 線維芽細胞培養におけるストレス[j]。^ free radic res,2004,38 (6): 601-611。
[31] cirillo n, vicidomini a, mccullough m,et al.ヒアルロン酸ベースの化合物は、酸化によって誘導される線維芽細胞老化を阻害します in vitroでのストレスとin vivoでの口腔粘膜炎の予防[j]。j cell physi - ol,2015,230(7):1421 -1429。
[32] Maytinエブヒアルロン酸:単なるシワ充填剤以上[j]。Glyco -生物学、2016年、26、(6):553-559。
[33] 赤ワイン煮 LY、Bikle D.Selective hyaluronan-CD44 signaling mirna-21の発現を促進し、ビタミンd funcと相互作用する。皮膚扁平上皮がんは紫外線照射後に進行する[j]。2015年Immunol前、6:224ました
[34] bourguignon ly, wong g, xia w,et (hyalu- ronan)相互作用 CD44 and RhoGTPase signaling promotes ケラチン生成細胞 functions and 克服 老化によって引き起こされる epidermal dys机能か[J]。j dermatol sci,2013,72(1):32-44。
[35] Lagatta A, D'アゴスティーノa, schiraldi c,et al. a biophysically-defined hyaluronic 酸ベースの化合物は、移行を加速し、 stimulates the production of keratinocyte-derived 神経調節物質[J]。cell adh migr,2019,13(1):23-32。
[36] lee sg, yoon ms, kim dh,et al Rosacea-Like ね through 消炎 表皮バリア改善効果[j]。^『仙台市史』通史編2(通史3)、189 -196頁。
[37] hu l, nomura s, sato y,et al. uvb誘発カルプロ・テクチン介在性ケラチノサイト炎症に対する異なる誘導性分子量ヒアルロン酸の抗炎症作用[j / ol]。J索引(Sci) Dermatol 2022.[2022-06-10]。 https: / www。 jdsjournal。 com e / article / s0923-1811 (22) 00158- x /fulltext。
[38] david-raoudi m, tranchepain f, deschrevel b,et al hyaluronan and its 線維芽細胞の断片:創傷治癒との関係[j]。傷 修理 2008年Regen、16 (2):274 - 287.
[39] ciccone v, zazzetta m, morbidelli l .血管新生に関連する線維芽細胞および内皮細胞機能に対する2つのヒアルロン酸製剤の効果の比較[j]。2019年細胞(1479 8(12):。
[40] asparuhova mb, kiryak d, eliezer m,et al.ヒト口腔線維芽細胞に対する2種類のhyalu- ronan製剤の活性[j]。^ a b c d e f g h i j pero,2019,54(1):33-45。
[41] quan t, wang f, shao y,et dermal microenvironment activates ヒトの高齢皮膚in vivoにおける線維芽細胞、内皮細胞、およびケラチノサイト[j]。^ a b c d e f g h i j invest dermatol,2013,133(3):658-667。
[42] Radrezza S男爵 G、Nukala SBメトロ・カードとデビッド al.Advanced quantitative proteomics to 評価 molecular effects of ヒト皮膚線維芽細胞における低分子ヒアルロン酸[j]。J Pharm肛門出来る20/20、185:113199。
[43] loren ce, dahl cp, do l,et al hyaluronan in human だぶだぶ 心筋症[J]。細胞 、2019年8(2):97に供給する。
[44] olsson m, bremer l, aulin c,et al alarmin function 評価 in 関節炎 synovial線維芽細胞 軟骨細胞と培養 文化[J]。 生まれつき Immun、 2018, 24 約数の和は131 -141。
[45] zheng bw, wang by, xiao wl,et al. dnvb誘導マウスアトピー性皮膚炎およびlps誘導生264.7細胞において、異なる分子量ヒアルロン酸が炎症反応を緩和する[j]。人生Sci、2022年301:120591。
[46] zhou j, ling j, wang y,et al.メラニン生成におけるインターフェロン-gam- maとインターロイキン-18のクロストーク[j]。J Photochem Photobiol B、2016年、163:133 -143。
[47] D'agostino a, la gatta a, stellavato a,et al. bioferの可能性-メンタティブな硫酸化されていないコンドロイチンと皮膚reペアにおけるヒアルロン酸[j]。^ int j mol sci,2022,23(3):1686。




