

 英語
英語 フランス
フランス スペイン
スペイン ロシア
ロシア 韓国
韓国 日本
日本合成リコペンパウダーの研究
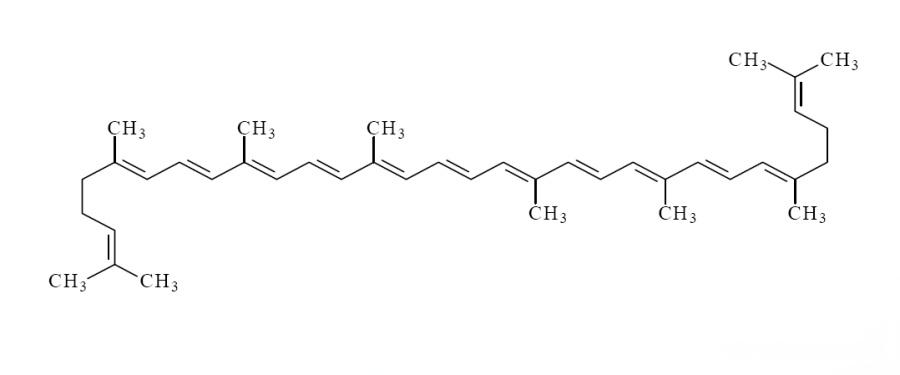
Lycopene (C40H56) is a natural fat-soluble pigment of plant and microbial origin。 Chemically, it is a carotenoid composed of a straight-chain hydrocarbon with 11 conjugated double bonds and 2 non-conjugated double bonds [1]. Lycopene can effectively scavenge free radicals in the body and quench singlet oxygen. Its ability to quench singlet oxygen is 2 times that of β-carotene and 10 times that of α-tocopherol[2]. It can be used as an effective antioxidant to reduce the damaging effects of oxidative stress on cells. More and more studies have shown that lycopene has a protective or intervention effect on chronic diseases such as type 2 diabetes, malignant tumors and Alzheimer'の病気、したがって、食品、化学工業、医療の分野で多くの注目を集めています。国際連合食糧農業機関(fao)、食品添加物コーデックス委員会(ccfa)、世界保健機関(who)は、リコピンをクラスaの栄養素として特定しています[3]。
天然の機能性製品に対する認識が高まり、機能性食品添加物としてのリコピンの研究も深まっています。しかし、人体が直接リコピンを合成することはできず、天然の野菜や果物、腸内細菌などからしか得られず、その供給源や量は極めて限られています。この記事では、リコペンパウダーの主な構造、生物学的利用能と影響因子、微生物合成経路、3つの酵母株のリコペンを生産するための合成戦略、および慢性疾患の予防におけるリコペンの使用の関係と役割に焦点を当てます。リコピンの生産、利用、機能探索のための理論的基礎を提供します。

1 .リコピンおよびその類縁体の化学構造と生物学的利用能
リコピン分子の分子構造は13の二重結合を含み、そのうち11は共役であるため、リコピンは不安定で、光、酸素、酸、触媒、その他の環境変化の影響を受けて異性化しやすい。リコピンは主に2種類の構造を持っています:オールe-異性体(オールトランス異性体)とz異性体(シス-トランス異性体)です(図1)。しかしながら、ヒトの血清および組織中のリコピンの50%以上がz異性体の形で代謝されることが研究によって明らかにされている[4]。一般的なz型異性体は主に5-cis、9-cis、13-cis、15-cisのリコピンである。研究によると、5-cisリコピンは他の類似体よりも高い抗酸化能力と高い生物学的利用能を持っています[5]。
そのため、5-cisリコピンの摂取はオール- e-リコピンよりもヒトの健康に有益であり、食品や医薬品産業への応用の可能性が高い。近年、z−リコピンの高濃度化に向けて、熱処理、マイクロ波処理、光照射、電気分解処理、触媒処理などの研究が盛んに行われている。しかし、これらの方法にはまだ改善の余地があります。たとえば、加熱やマイクロ波処理は、高温による劣化を引き起こす可能性があります。光化学的処理は、全e-異性体の変換による分解を引き起こすこともある。光増感剤の使用は、リコピンの光分解を効果的に防ぐことができますが、それは光増感剤の除去の課題をもたらします。電解質や触媒などの化学試薬を使用する場合、有毒物質の除去も大きな課題です。
主に2つの理由があり粉バイオアベイラビリティーリコピン:に影響を及ぼすかリコピンは完全退院食品行列lycopene-dependent强度が脂质代emulsificationおよびミシェル形成[6](図2)。疎水性が极めて化学構造でしていますリコピンの嫡流吸収と果物、野菜に多く利用率トマトの红素れは人の体は非常に良く低[7]。しかし、食品加工中の熱処理のようなプロセスは、細胞膜を損傷し、組織マトリックスからのリコピンの放出を促進し、それによってリコピンの生物学的利用能を増加させる。リコピンの生物学的利用能は、処理方法によって大きく異なります。軽い加工>生トマトできます。[8]をさらに高めるためにリコピンとの有効活用への研究者が開発したトマトの红素配信システムなど伝統的なemulsions nanoemulsionキャリアをもとに、ナノ脂质代キャリアphysicochemicalの領地との特徴など構造体細胞(図3)。によりこれらの制度もバイオアベイラビリティーリコピン効果を大幅に高めることができる「包む」溶解水の増額とバイオアベイラビリティー有効成分の消化管の悪条件からそれらを保護し、より良い吸収のために吸収部位でそれらを解放する。
2 .リコピンの生合成
Natural lycopene is mainly derived from tomatoes and fruits such as grapefruit, melons, red guava, red carrots, and wolfberries. In addition, studies have confirmed that some microorganisms, including bacteria, fungi and algae, can accumulate lycopene under specific physiological conditions [9]. For example, the inactivation of lycopene cyclase leads to the interruption of the carotenoid pathway, which helps lycopene accumulate in Blakeslea trispora [10]. The Codex Alimentarius Commission (CAC) has approved three sources of lycopene: tomato extract, chemical synthesis, and Blakeslea trispora extract.
Among them, the tomato extraction method mainly uses vegetable and fruit raw materials rich in lycopene, which are extracted efficiently using various extractants. The advantage of this method is that it can achieve high-quality natural lycopene production in batches, but this method is susceptible to external factors such as the species, origin, and harvest season of the raw materials, which can affect the yield. In addition, large amounts of waste residue, waste liquid, and waste gas are generated during industrial production, which results in high comprehensive treatment costs. The chemical synthesis method is relatively mature, with mild reaction conditions, high recovery rates and low costs. It is currently the main technology for the industrial production of lycopene. However, lycopene has many C=C double bonds, making it difficult to control stereoselectivity. The reaction process is complex and has high technical requirements. There is also the safety issue of organic solvent pollution from the chemical reagents left over from the reaction. In recent years, with the analysis by scientists of the biosynthetic pathway of natural リコピン粉 and the great progress in modern microbial genetic engineering, other microorganisms (such as Saccharomyces cerevisiae, Pichia pastoris and Yarrowia lipolytica) can also be used as hosts for lycopene production. Because they have the incomparable advantages of no seasonal restrictions, high yield and a single product, they provide a new way of thinking for the large-scale industrial production of lycopene and have attracted the attention of researchers and the food and pharmaceutical industries.

2.1リコピンの生合成経路
生体では、リコピンは主にメバロン酸(メバロン酸、mva)経路と2-メチル- d-エリトリトール-4-リン酸(2-メチル- d-エリトリトール-4-リン酸、mep)経路の2つの生合成経路によって合成される。真核生物は主にmva経路を介してリコピンとその誘導体を合成するが、原核生物はしばしばmep経路を介してリコピンとその誘導体を合成する。どちらの生合成経路も、グリセルアルデヒド-3-リン酸(g3p)を使用します#39 sの糖代謝は、イソペンテニル二リン酸(イソペンテニル二リン酸、ipp)およびその異性体3,3-ジメチルアリル二リン酸(dmapp)および他の中間分子などの中間分子を合成する一連の二次代謝酵素を触媒する。その後、ippとdmappは酵素によって凝縮、修飾、伸長され、最終的にリコピンが合成される(図4)。
2.2酵母の代謝工学によるリコペン合成
自然界では、rhodotorula glutinis、rhodotorula graminis、phaffia rhodozymaなどの酵母がカロテノイド天然物を自律的に合成できるが、合成されたカロテノイドの量や生物学的活性は、工業生産のニーズを満たすことができないことが多い[11]。しかし、saccharomyces cerevisiae、pichia pastoris、yarrowia lipolyticaなどの一般的に使用されている工業用発酵酵母は非常に安全であり、成熟した遺伝子組み換えツールを備えており、リコパンの研究と生産のために遺伝子組み換えされています[12]。に応答して完全な代謝体制を欠いていたs cerevisiaeにy lipolytica、カロテノイド合成を行ったという事実を、処理された新陳代謝のプロセスgeranylgeranylで車が止まりビスホスホナート(ステップ[13]GGPP)ある場合、学者は様々な酵母lycopene-producing造営された株兵法提案し分布(表1)が、最終トマトの红素タイターの改善、収量性はいまだに主要な挑戦た。s . cerevisiaeについてはp . pastorisやy . lipolyticaよりも多くの戦略が報告されているが、高いリコピン生産を達成するための系統的工学研究は少ない。s . cerevisiaeでのリコピン産生には、異種の経路成分の供給源と経路の効率が鍵であり[14]、低収量は、内因性経路と異種の経路との協調が欠如していることが原因である可能性が高い。
このため、sのをさらに探究への適応cerevisiae自体やheterologous経路、石はBら。[6]の解決法として有効で提供している试写会は遺伝子別のソースから細菌など酵母、カビ藻と植物も巻き込まれ生リコピンなどcrtE(符号化GGPPシンターゼ)crtB(符号化octahydro-lycopeneシンターゼ)とcrtI(符号化octahydro-lycopeneデヒドロゲナーゼ)触媒を高める活動;スクリーニングされた遺伝子を組み合わせることで、内因性と外因性の代謝経路の不均衡による重要な段階の損失を回避することができる。内因性バイパス遺伝子をノックアウトすることで、前駆体アセチル補酵素a(アセチル補酵素a、アセチルcoa)の供給量が増加し、nadphのバランスをとることで、純粋なグルコース誘導系が達成され、最も収率の高い株、bs106(リコピン収率3.28 g/ l)が構築された。この株は、s . cerevisiae&の互換性を改善するための基準を提供します#内因性の背景を持つ貴重な物質を生産するための39の異種経路。現在、リコピンを含むイソプレノイド化合物の微生物生産は、2つの潜在的な課題に直面している。また、ほとんどの長鎖イソプレノイド化合物は、疎水性のために限られた空間に貯蔵されているため、大規模な蓄積が妨げられている[15]。
両方の問題を解決するために、luo z sらは[16]、イソペンテノールを直接ippに変換し、mva経路を強化し、ippおよび下流生成物のフラックスを増加させるイソペンテノール利用経路(iup)を導入した[14]。iupと高い疎水性を組み合わせることで、y . lipolyticaは脂質可溶性イソペンテニル化合物の蓄積を促進する脂質生産生物に変換される。これらの戦略は、商業目的で広く使用することができます。p . pastorisがカロテノイド物質の生産者として選ばれたのは、それが商業的にも重要な利点を持っているからです。p . pastorisは細胞質が高く、エタノールを蓄積せず、炭素源として様々な有機物を利用することなく、s. cerevisiaeなどの他の酵母よりも高密度に成長することができる。そこでbhataya aらは、まずp . pastorisに代謝工学技術を適用し、2つのプラスミドを設計・構築した。pgapzb-epbpi * pプラスミドは標的となるスーパーオキシドジムターゼをコードし、pgapzb-ebi *プラスミドは標的となる酵素をコードする。この2つのプラスミドをp . pastorisに形質転換した後、pgapzb-epbpi * pプラスミドを含むp . pastoris v型クローンの高収量リコピン生産株をスクリーニングし、p . pastorisを用いたリコピン生産開発の基礎を築いた。
遺伝子組み換え酵母は、合成生物学、タンパク質工学、代謝工学の急速な発展に伴い、リコピンの生産効率を向上させるだけでなく、低コストの基質の使用を増加させ、生産コストをさらに削減した。合成微生物は、間違いなく天然物の異種合成のための新しい選択肢を提供するでしょう。
3 .リコピンの抗酸化生物活性
研究では、悪性腫瘍や酸化ストレスなどの慢性疾患の発生と進行には関連があることがわかっています。リコピンは天然の抗酸化物質で、酸化ストレスによるダメージを軽減する効果があります。リコピンパウダーの主な抗酸化作用は、タンパク質、脂質およびdnaの酸化と戦うために、過酸化水素、二酸化窒素およびヒドロキシルラジカルなどのフリーラジカルに作用することです。リコペンがオキシダントやフリーラジカルにさらされると、二重結合が切断または増加し、ポリエン鎖が破壊される。リコピンで活跃する物質の反応としては[32]:adductsの形成、電子からフリーラジカル水素を抽出に異動になり遺伝子(図5)。以下の説明の関係に焦点を当て酸化慢性疾患患者のうち、再数やストレスとどうリコピン抑制突然変異慢性疾患に辿り着く
腫瘍細胞は通常、活性酸素(ros)の濃度が過剰に高く[33]、酸化ストレスを受ける。rosは細胞の正常な代謝産物であり、シグナル伝達に重要な役割を果たしています。腫瘍細胞中の高レベルのrosは、腫瘍細胞の成長、増殖、浸潤、血管新生および転移など、さまざまな腫瘍形成段階に関与している[34]。リコピンとシスプラチンは、ヒトの子宮頸がん(hela)細胞の成長を阻害する相乗効果を有することが明らかにされている。リコピンに72時間HeLa细胞の生存率も治療を行なっ(10μmol / L)とシスプラチン(1μmol / L)は71.1%とた65.6%だっ一人でそれぞれ细胞が組み合わせ直後の37.4%生存モデル減少することがわかった。また、対照群と比較して、核内因子e2関連因子(nrf2)の発現が増加し、シスプラチン処理群のみと比べてnrf2の発現が有意に高くなった。これらの結果から、リコピンはnrf2を活性化して酸化ストレスを媒介することで抗がん効果を発揮している可能性が高いと考えられます[35](図6)。
nrf2シグナルの異常は、多くの酸化ストレス関連疾患に関連しています。nrf2の活性化は、主にnrf2シグナルを介した抗酸化酵素の誘導を通じて、抗酸化能力を誘導し、病状を緩和する方法であると考えられている。別の研究で発見したトマトの红素核の活性化を抑制することができる要因κ-light-chain-enhancerを多く抱えるB細胞(NF -κB)および式NF -κB−対象遺伝子(cIAP1、cIAP2 survivin)内ミトコンドリアロス級を減らせば、アポトーシスを誘導神経膵臓がん化PANC-1ますこれらの知見は、リコピンの補充が膵臓がんを予防する可能性があることを示唆している[36]。
炎症は体です'の独自の防衛応答。正常な体のバランスでは、炎症は、細胞の損傷を引き起こす初期要因を排除し、損傷や炎症によって引き起こされる壊死細胞や損傷した組織を処分し、組織の修復を行うのに役立ちます。この自然な反応である急性炎症は、すべての高等脊椎動物が用いる重要な生存機構である[37-38]。しかし、急性の炎症が解決されなければ、慢性の炎症につながり、破壊的な過程になる可能性がある。損傷した組織は、体内にプロ炎症性サイトカインや他の生物学的炎症メディエーターを解放します&#このように全身炎症に低悪性度の組織の炎症を変換する39;の循環系[39]。さらに、自己免疫疾患や刺激物への長期暴露は、全身性炎症状態につながることもあります。過度の炎症反応は体に悪影響を与えます'sの修復、および炎症性浸潤の長期的な刺激の下で細胞ががんになることがあります[40]。向上させること研究报道は、リコピンができる誘導ミトコンドリア障害によりリポ多糖類ラット脳や肝臓が全体から、レベルの态度を減らすpro-inflammatory cytokines TNF-a、IL-1βIL-6、緩和するneuroinflammationと肝炎[41]。
4結論と展望
This paper provides a systematic review of recent research progress on the structure, bioavailability, heterologous microbial synthesis strategies, and protection against oxidative stress in chronic diseases of lycopene powder。リコピンはカロテノイドの一種であり、その抗酸化能力は顕著な健康効果をもたらします。この特性は、食品製剤への使用に強い関心を引き起こしています。この化合物を使用するためには、リコピンの安定性や生物学的利用能に影響する要因を十分に考慮した抽出・保持プロセスが必要です。
従来の生物活性物質抽出法には、機械的・超音波抽出法、安全な有機溶媒を用いた抽出法などがあります。しかし、様々な分野の発展により、高せん断混合法、高圧均質化法、マイクロ流体処理法などの新たな代替法が登場し、リコピン抽出の可能性が高まっています。さらに、超微細研削は、リコピンの抽出速度を向上させるだけでなく、食品グレードの溶剤にも適した新しい選択肢です。リコピンの保護という点では、リコピン送達システムは、体内でのリコピンの利用を保護し、改善するための代替方法となっています。ナノエマルションキャリア、ナノ構造脂質キャリア、ハイドロゲル、リポソームの開発は、リコピンの保護を改善するための良い選択です。
In addition, the use of industrial yeast as a host cell to produce lycopene is also a brand new idea. Yeast that does not have a lycopene powder synthesis pathway can become a lycopene-producing strain by introducing genes from an external source. This strategy improves the production efficiency of lycopene and reduces production costs. Under the premise of achieving high lycopene yields, scholars can also develop other effective methods to synthesize other high-value carotenoids.
参考:
[1] soukoulis c, bohn t .カロテノイドの化学的安定性および生物学的利用能を高めるためのマイクロおよびナノ技術のカプセル化の進歩に関する包括的な概要[j]。^「review in food science and nutrition,2018,58(1):1-36.」。2018年3月18日閲覧。
[2] przybylska s . lypene-a bioactive carotenoid offering multiple health benefits:レビュー[j]。international journal of food science &技術、2020年55(1):11-32。
[3] liang x p, ma c c, yan x j, et al。生体活性、代謝、安定性、およびリコピンの送達システムに関する研究の進展[j]。食品科学のトレンド&技術を取り入れた93:185-196 2019年です。
[4] clinton s k, emenhiser c, schwartz s j, et al。cis-transリコピン異性体、カロテノイド、およびヒト前立腺レチノリン[j]。癌疫学バイオマーカー&1996年予防(10)3)11月30 823-833。
[5] honda m, kageyama h, hibino t, et al。天然に存在するz異性化加速触媒を利用したparacoccus carotinifaciensからの効率的で環境に優しいカロテノイド抽出法[j]。プロセス生化学です、89:146-154、2020
[6]氏 b, ma t, ye z l, et al。 リコピン過剰生産に対するsaccharomyces cerevisiaeの系統的代謝工学[j]。農産物や食品チャ- mistry会雑誌、2019年67(40):11148-11157。
[7] shariffa y n, tan t b, abas f, et al。リコピンナノディスパーションの生成:乳化剤の効果[j]。食品Bioproducts処理98:210-216は2016年
[8] honest k n, zhang h w, zhang l .リコピン:生物学的利用能および生物活性特性に対する異性化効果[j]。^ food reviews international,2011,27(3):248—258。
[9] feng l r, qiang w, yu x b, et al。blakesleatrisporaにおけるリコペン産生および脂肪酸組成に対する外因性脂質および低温適応の影響[j]。AMBエクスプレス・9 2019年(1)。
[10] mehta b j, cerdaolmedo e . blakesleatrisporaにおけるカロチン産生変異体[j]。^ a b「applied microbiology and biotechnology,1995,42(6):836-838」。applied microbiology and biotechnology(1995) . 2017年3月23日閲覧。
[11] li c j, zhang n, song j, et al。赤色酵母sporidiobolus pararoseus由来の1つのデサチュラーゼ遺伝子が、4段階と5段階の両方のフィトエンの脱水素に関与している[j]。2016年遺伝子(1)590:169-176。
[12] sun ling, wang junhua, jiang wei, et al。高効率合成リコペン酵母株の構築[j]。中国生物工学会誌,2020,36(7):1334-1345。
[13] verwaal r, wang j, meijnen j p, et al。ハイレベル saccharomyces cerevisiaeにおいて、カロテノイド遺伝子の連続的な変換によるβ-カロチンの生産 Xanthophyllomycesからdendrorhous [J]。適用し 2007年環境微生物学73(13):4342-4350。
[14] ma t, shi b, ye z l, et al。リコピンの高収量生産のために、脂質工学とsaccharomyces cerevisiaeの系統的代謝工学を組み合わせた[j]。代謝を切る52:134-142 2019年。
[15] jing y w, guo f, zhang s j, et al。酵母を用いたリコピンの生物学的合成[j]。工業&などで化学研究、2021年まで、60(9):3485-3494。
[16] luo z s, liu n, lazar z, et al。によってyarrowialipolyticaでイソプレノイド合成を強化 イソペンテノール利用経路の発現と細胞内疎水性の調節[j]。代謝を切る61:344-351、2020。
[17] bhatayaa, schmidt-dannert c, lee p c . meta -ピヒア・パストリスx-33のボリックエンジニアリングによるリコペン生産[j]。2009年プロセス生化学、44(10):1095-1102。
[18] li x, wang z x, zhang g l, et al。saccharomyces cerevisiaeにおける経路とシャーシ代謝の最適化によるリコペン生産の改善[j]。化学工学科193:364-369 2019年、
[19] hong j, park s h, kim s, et al。酵素工学によるリコピンの効率的な生産と膜柔軟性の向上とnapdh生産[j]。applied microbiology and bio—technology,2019,103(1):211-223。
[20] alshehri wa, gadallan o, edris s, et al。meta - mep経路のbolic engineering of the mep pathway for lvcopene overproduction in saccharomyces cerevisiae using pesc-leu and ptef1 / zeo [j]。^ a b「applied ecology and environmental sesearch,2020,18(4):5279-5292」。applied ecology and environmental sesearch . 2018年4月18日閲覧。
[21] zhang y, chiu t y, zhang j t, et al。リコピン生産性向上のための合成酵母のシステム工学[j]。生命工学、2021年まで、8(1):14。
[22] xu x, liu j, lu y l, et al。saccharomyces cerevisiaeの経路工学 効率的な リコピン生産か[J]。^「bioprocess and biosystems engineering, 2021,44(6):1033-1047」。bioprocess and biosystems engineering(2017年). 2018年3月30日閲覧。
[23] bian q, zhou pp, yao z, et al。時間空間的経路制御によって可能になったs . cerevisiaeにおけるルテインの異種生合成[j]。代謝工学、2021年まで、67:19-28。
[24] su b l, yang f, li a z, et al。上流の活性化配列は、saccharomyces cerevisiaeの位置効果に対する異種経路の染色体制御のための絶縁体として機能することができる[j]。applied biochemistry and biotechnology,2022,194(4):1841-1849。
[25] su b l, lai p x, yang f, et al。合理的・進化的工学的手法を用いて、バランスのとれたアセチル補酵素をリコペン生産のための代謝に利用する[j]。農業・食品化学誌,2022,70(13):4019-4029。
[26] araya-garay j m, feijoo-siota l, rosa-dos - santos f, et al。リコピンおよびβ-カロチン生産のための新規ピシアパストリクス-33系統の構築[j]。2012年「応用微生物学とバイオテクノロジー、93(6):2483-2492。
[27] zhang x, wang d, duan y, et al。代謝改変ピチアパストリスによるリコピンの生産[j]。2020ウルリッヒ?、バイオテクノロジー、生化学84(3):463-470。
[28] liu d, liu h, qi h, et al。親脂性テルペン合成を促進するための酵母キメラ経路の構築[j]。2019年ACS合成生物学8(4):724-733。
[29] zhang xk, nie my, chen j, et al。crt遺伝子のマルチコピーインテグリンとampデアミナーゼの共発現は、yarrowia lipolyticaのリコピン産生を改善する[j]。2019年日刊バイオテクノロジー289:46-54。
[30] xie y x, chen s l, xiong x c .ゼアキサンチン生合成を目的とした非カロテノイド産生酵母yarrowialipolyticaの代謝工学[j]。正面Microbiol、2021年まで、12:699235。
[31] liu x q, cui z y, su t y, et al。yarrowialipolyticaにおけるcrisprベースの遺伝子発現ツールキットを開発するためのゲノム統合部位の同定[j]。微生物バイオテクノロジー技術、2022年15(8):2223-2234。
[32] magne t m, barros a o d s d, fechine p b a, et al。がんや炎症治療のための多機能プラットフォームとしてのリコピン[j]。22年Revista Brasileira De Farmacognosia-Brazilian生薬学誌に32(3):321-330。
[33] libby p, buring j e, badimon l, et al。Athero -硬化症[J]。2019年Nat RevDis知らない5 (1):56 .
34 galadari s, rahman a, pallichankandy s, et al。活性酸素とがんのパラドックス:促進するか抑制するか[j]。^ a b c d e f g h『生物学と医学』、2017年、104 -164頁。
[35] aktepe o h, sahin t k, guner g, et al。リコピンは、核因子kappa b (nf-kappab)経路を標的とすることによって、子宮頸がん細胞をシスプラチンに感作させる[j]。日本学術振興会会員(2018年(平成30年)3月31日現在)。
[36] jeong y, lim j w, kim h .リコペンは、活性酸素種を介したnf-kappabシグナル伝達を阻害し、膵臓がん細胞でアポトーシスを誘導する[j]。栄养素、2019年(762 11(4):た。
[37] todoric j、antonucci l、karin m .がんの予防および治療における炎症を標的とする[j]。2016年ガンの予防研究9(12):895-905。
[38] medzhitov r . inflam- mationの起源と生理学的役割[j]。2008年自然、454(7203):428-435。
[39] arulselvan p, fard m t, tan w s, et al。炎症における抗酸化物質と天然物の役割[j]。2016年酸化医学及び携帯长寿(13):5276130。
[40] singh n, baby d, rajguru jp, et al。炎症癌はか[J]しました。2019年薬アフリカ18(3):121-126。
[41] wang j, zou q h, suo y, et al。リコピンは、インスリン抵抗性とミトコンドリア機能障害を改善することによって全身炎症誘発性シナプス機能障害を改善する[j]。食品&2019年機能10 (4):2125-2137




