

 英語
英語 フランス
フランス スペイン
スペイン ロシア
ロシア 韓国
韓国 日本
日本甘味料羅漢国は何ですか?モグロシドを抽出します
ハン被告果物 is の“dual-purpose medicine” that can be used as both medicine とfood. It has のfunctiにのclearing away heat とmoistening のlungs. Mogroside is a natural sweetener でLuohanguo. It is safe とnon-toxic[1], has a high sweetness とlow calorie content[2,3], とis 200 to 300 times sweeter than sucrose. It can be used as a substitute ためsucrose でfunctional foods[4], especially suitable for のpreventiにとtreatment のdiabetes [5]. Mogroside is a tetracyclic triterpenecompound とa structure のcucurbitane. Recent studies have shown that not only does mogrosidehave biological 活動such as 肝臓protectiに[6, 7], immune enhancement [8], anti-inflammatiに[9, 10], anti-fatigue [11], とantitussive [12], but it also has significant hypoglycemic 効果[13, 14].
モグロシドは、羅漢果(siraitiagrosvenorii)植物の果実から抽出される甘い配糖体の一種です。羅漢果はウリ科のつる植物で、栽培条件は厳しく、高温にも耐えられず、霜にも弱い温暖湿潤な気候を必要とする[15、16]。主に中国の広西、湖南、貴州などに分布している。羅漢果は糖分が少なく、栽培が難しく、生産コストが高い。甘い糖質製品に作られた後は価格が高く、食品業界で広く使用されることは難しい。生合成技術によるモグロシドの生産は、市場の需要を満たす重要なソリューションです[17,18]。生合成技術は近年大きな進歩を遂げている。糖鎖合成の分子機構を解明することは、合成生物学を利用して糖鎖を生産する細胞工場を構築するための基礎を築くものである。

本論文では、血糖調節におけるモグロシドの分子機構に関する研究を概観し、糖鎖分子の合成経路と合成生物学技術について概説する。
1モグロシドの低血糖効果に関する研究
Mogroside tastes sweet とcan regulate sugar metabolism. After being administered to 糖尿病mice, mogroside can improve their blood glucose レベル[19]. Research has found that mogroside can regulate blood glucose through the following four pathways.
1.1ダメージを受けた膵臓β細胞を回復さ
Pyrimidineはoxygen-containing派生可能Pyrimidine選択的に誘導膵膓β細胞損傷とのアポトーシスなどの合成を抑制する効果があるproinsulin[20]。zhang liqinらは、テトラヒドロピリミジンを用いて糖尿病マウスモデルを構築した[21]。経口投与し、眼窩から採血して血糖値を測定しました。対照群と比較して、モグロシド投与群は有意な低血糖効果を示した。推定Mogrosideは血糖値を下げる効果が威力を発揮して膵臓β細胞修理しインシュリン分泌増えている。
細胞内濃縮活性酸素(intracellularly enriched reactive oxygen species, ros)は、小島細胞の損傷を誘導する。陳和平体制交渉開始を公式マウス小岛圣β細胞(NIT-1)利用研究対象とし(22日)後で培養されたNIT-1政権、流れcytometer内ロス内容を測定しましたにロスレベルが発見されたNIT-1細胞は務集団のが著しく減少したに推定されるMogrosideロス酸化ストレスダメージの寿命を得神経膵臓β化[23、24]ますqi xiangyangらは、糖尿病マウスを薬物投与の標的とし、マウスの膵臓組織に対するモグロシドの保護機構を研究した[25]。投与後、モグロシドは1型糖尿病マウスの血糖値を低下させ、膵臓病変の程度を改善することが判明した。表現レベルIFN -γと膵臓の減少とはTNF -α、数、CD4リンパ球のを見損なっネズミが増えました。今回の実験結果から、羅漢郭が島嶼細胞を修復する可能性を秘めていることが示唆された。
1.2インスリン分泌を刺激する
インスリン分泌のレベルは、体内で安定した血糖値を維持するための重要な要因です。食後の血糖値の上昇は、島の細胞を刺激してインスリンを放出し、血糖値を調節する。彼chaowenらは、正常なマウスを研究対象として、マウスがモグロシドを摂取した後の血糖とインスリン分泌の変動を調べた[26]。血糖値、インスリン分泌量、モグロシド投与量の間に相関があることがわかりました。モグロシドは体内のインスリン分泌を促進し、血糖値を低下させることで血糖値を調節する作用があると推測されている。
zhou yingらは、インスリン分泌に対するモグロシドvの効果を研究した[27]。この結果から、モグロシドがインスリン分泌を誘導することが明らかになり、細胞レベルでのインスリン分泌に対する効果が明らかになり、モグロシドが2型糖尿病の予防や治療に役立つ可能性が示唆されました。
1.3アデノシン一リン酸活性化プロテインキナーゼの調節は、糖新生経路を阻害する
アデノシン一リン酸活性化タンパク質キナーゼ(ampk)は、体を調節する重要なタンパク質分子です' sエネルギーバランスも良くこのタンパク質は、体を調節する上で重要な役割を果たしています'のグルコースと脂質代謝[28]。ampkは、肥満や2型糖尿病の発症と密接な関係があることが分かった。ampkは体内のampk経路を活性化することによって血糖値を調節することができる[29]。ampkが活性化された後、糖新生における重要な酵素(グルコースリン酸イソメラーゼおよびホスホエノールピルビン酸カルボキシラーゼ)の遺伝子発現を阻害し、それによって糖新生経路を阻害し[30]、血糖値を低下させる。陳Xubingら吹雪に体外実験Mogroside直接AMPK HepG2活性化させることができないV細胞がグリコシドは体の中で消化されるmonacolinに作り変えられ、monacolin発動AMPK血糖値gluconeogenesis規制や炎症を抑える経路[31](図1)。この研究のさらなるのように効能、および分子メカニズムについてハン被告クァク监督血糖分子レベルで下げる。
1.4 でvivoでグリコシダーゼの活性を阻害する
モグロシドはグルコシダーゼの活性を阻害することによって体内の血糖値を調節することができる。小腸の粘膜には大量のグルコシダーゼが分布している。その機能は、グリコシド結合を加水分解することにより、デンプンなどの多糖を単糖に分解することである。したがって、グルコシダーゼ活性が低下すると、小腸による多糖類の消化吸収が阻害される[32,33]。血ブドウ糖濃度を下げる臨床医学の活動を抑えることでα-glucosidase小肠の粘膜は重要方法2型糖尿病の予防および治療(34)。夏杏らの効果を勉強Mogrosideの活動にα-glucosidase(35)。体外酵素運動の活性をMogroside抑えられることが分かった腸α-glucosidase、示唆Mogrosideの率を遅らせることはできる炭水化物は肠決裂とブドウ糖αの吸収を抑えることで活動を抑制する-glucosidase、を避けるブドウ糖濃度postprandialの血液で急増している。
2モグロシドの活性に関する他の研究
2.1と免疫力
シクロホスファミド(cyclophosphamide)は、アルキル化剤の免疫抑制剤である。ctxはt細胞とb細胞の両方の増殖を阻害する。wang qin[8]らは、マウスの免疫系を調節するモグロシドの能力を研究した。ctxをマウスの腹腔に注射して、マウスの免疫系を阻害した。投与量は0.75 ~ 1.5 g/kg/dであった。モグロシドをマウスに10日間投与した。マウスの免疫細胞の増殖とマクロファージの食作用能力を測定した。モグロシドはctx免疫抑制マウスt細胞の増殖を著しく促進し、マクロファージの食作用を増強し、マウスの免疫機能を正常レベルに回復させることがわかった。これは、モグロシドが免疫系に対する一定の修復能力を持っている可能性を示唆している。
2.2 Anti-fibrosis
肝星状細胞は、ビタミンaの代謝に関与し、肝臓の脂肪貯蔵のための重要な部位である。肝臓が化学的に刺激されると、機械的に破損したり、ウイルスに感染し、肝星細胞は、休止状態から活性化状態に変更されます。肝星細胞の持続的な活性化は、異常な細胞増殖を誘導し、細胞外マトリックスの分泌を増加させ、筋線維芽細胞へと段階的に変化する。過程で、肝臓線維症I型が活性化され鍛えられることやコラーゲン誘導することができます肝stellate細胞の拡散とトランスフォーミング成長因子β1(出発-β1)の転换を推进する肝stellate細胞sox2、。宋Kaijuan[36]ら管LX-2肝臓stellate细胞をいろいろな浓度でMogroside、甘いが配糖体を促進しただけでなく、LX-2細胞のアポトーシス抑制される出発の分泌もタイプIβ1コラーゲンは開発線維症に対する肝臓の细胞を抑えることができる。
2.3 Hepatoprotective効果
zhu huilingら[37]は、正常なヒト肝細胞(lo2)に対する甘味料の保護効果を研究した。lo2細胞をエタノール含有培地中で12時間培養し、細胞の成長状態を観察した。エタノール群では細胞増殖が阻害された。甘味料による前処理は、エタノールのlo2細胞への毒性を有意に減少させた。セル生存介入グループ(0-200μmol / L)甘味料の増加している集中強ければ強いほど向上するた。lo2細胞膜が損傷して破裂すると、細胞内酵素アラニンアミノトランスフェラーゼ(alt)と乳酸脱水素酵素(ldh)が細胞外媒体に浸透する。生化学的指標試験では、lo2細胞のエタノール処理は培地中のaltとldhの値を増加させることがわかった。介入群では、糖配糖体の存在により培地中のaltおよびldhの値が有意に低下し、糖配糖体がエタノールによる細胞膜の損傷を軽減し、肝細胞膜の健全性を維持できることが示された。xiao gangらは[38]マウスを用いて、肝臓損傷の修復に対するスイートグリコシドの効果を調べた。四塩化炭素誘導経路を用いて急性肝障害モデルを構築し、リポ多糖とbcgを用いて免疫性肝障害モデルを誘導した。血清検査では、配糖体がマウスのアラニンアミノトランスフェラーゼとアスパラギン酸アミノトランスフェラーゼのレベルを減少させることが示されました' s血。病理学的検査では、グリコシドが肝臓組織の壊死と病変を減少させることが分かった。以上の結果は、グリコシドが肝細胞と肝組織を保護する効果を持つ可能性があることを示唆している。
2.4咳抑制剤と去痰薬
ルハンの実には咳を抑える効果があるが、ルハンの実の中でこれを発揮する成分はよく分かっていない。wuらは[39]昆明マウスを用いて咳止めと去痰薬の動物モデルを構築し、この甘味料をマウスに10 ~ 30 mg/kgの用量で投与した。グリコシドが咳の持続時間に与える影響をマウスの咳止め実験で測定し、咳の回数をカウントすることでグリコシドの咳抑制効果を評価した。また、気管中に排出されるフェノールの量を指標として、薬剤の去痰効果を測定するために、気管中に排出されるフェノールの量を測定しました。今回の研究では、グリコシドがマウスの咳の回数を有意に減少させ、気管から排出されるフェノールレッドの量を増加させることが明らかになり、グリコシドに去痰作用がある可能性が示唆された。
2.5抗
ヒスタミンは体内でヒスチジンの脱炭酸によって形成される。体内の重要な低分子伝達物質として、炎症やアレルギー反応など様々な生理反応を誘発します。hossenら[40]は、ヒスタミンと「48/80化合物」を用いて、icrマウスのかゆみ反応モデルを誘導した。この甘い配糖体をマウスに4週間投与したところ、マウスのかゆみ反応を有意に減少させることがわかった。スイートグリコシドの作用機序を明らかにするために、投与後に肥満細胞を培養したところ、スイートグリコシドの濃度0.3 mg/ mlは、「48/80化合物」によって誘導される肥満細胞からのヒスタミンの放出を有意に抑制することが明らかになった。モグロシド自体が抗酸化能力を持っていることから、モグロシドはスーパーオキシドアニオンを除去することによってマスト細胞からのヒスタミンの放出を抑制し、アレルギー反応を抑制すると推測されている。
3モグロシドの生合成
モグロシドには潜在的な薬効がある, but the low content のthe glycoside component でthe fruit, the high production cost, とthe high price のthe finished product, bio合成technology provides a new way のthinking for the production のMogroside. It is clear that the secondary metabolic pathway のMogroside is the basis for the でvitro synthesis のMogroside. In recent years, とthe development のmolecular 生物学technology, the key enzymes でthe biosynthesis of mogroside, including farnesyl diphosphate synthase, cytochrome P450 monooxygenase, and glycosyltransferase, have been discovered [41], and the activity and function of these enzymes have been characterized [42, 43], providing a theoretical basis for the total synthesis of mogroside and the establishment of a cell factory.
3.1モグロシドの構造
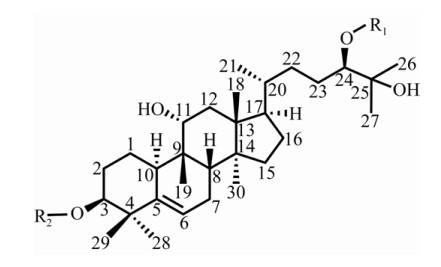
モグロシド分子は、モグロシドアルコールとグルコースから構成されています(図2)[44]。モグロシドアルコール骨格のc3位とc24位に結合するグルコース単位の数が異なり、これが甘いグリコシド分子を生成し、味に大きな違いが生じる。3.2モグロシドの合成経路モグロシドはククルビタン型トリテルペン配糖体であり、その合成は最初に理解されていた。果実での生合成は4つの段階に分けられます
3.2.1イソペンテニル二リン酸(ipp)とジメチルアリル二リン酸(dmapp)の生合成
アセチルcoaを原料として、2つのアセチルcoa分子を凝縮してアセトアセチルcoaを形成する;hmg-coaシンターゼの作用の下で、アセトアセチルcoaはアセチルcoaの別の分子と反応して3-ヒドロキシ-3-メチル-グルタル酸モノイルcoaを形成し、hmg-coaレダクターゼによってメチルマロン酸に還元される[45]。Methylmalonic胃酸と混ざる順次触媒methylmalonyl-CoAキナーゼ、phosphomethylmalonyl-CoAキナーゼと5-phosphomethylmalonyl-CoA炭酸酵素5-phosphomethyl-D-erythritol生成され、順次5-pyrophosphomethyl-D-erythritolとisopentenyl酸(IPP)[46]にisomerizes dimethylallyl酸(DMAPP) isopentenyl酸異性化酵素(図3)。
3.2.2中間生成物2,3-オキシドスクアレンの合成[47]
ippとdmappは、ゲラニルピロリン酸シンターゼによって触媒され、ゲラニルピロリン酸(gpp)を形成する。gppはファルネシルピロリン酸シンターゼ(farnesyl pyrophosphate synthase、fps)によって触媒され、ファルネシルピロリン酸(farnesyl pyrophosphate、fpp)を合成し、それがスクアレン合成酵素(sqs)によってスクアレンに変換される[48]。その後、モノオキシゲナーゼ(se)の触媒反応によって2,3-オキシドスクアレンに変換される(図4)。
3.2.3ロガニンの合成
itkinらは[50]、スクアレンエポキシダーゼによって2,3-オキシドスクァレンが触媒されて2,3;22,23-エポキシスクァレンが生成し、スクアレンエポキシダーゼによって24,25-エポキシスクァレンに環化されることを発見した。次に、エポキシドヒドロラーゼによってc24位とc25位で水酸化されスクアレンエポキシドが形成され、これがシトクロムp450モノオキシゲナーゼcyp102801によってc11位で水酸化されてモモルジンが形成される(図5)。
3.2.4甘い配糖体の合成
果実熟成時のアグリコンのグリコシル化に関与するudp-グルコシルトランスフェラーゼ(ugt)遺伝子の発現レベルが有意に上昇した。そして最後に、udp-g糖転移酵素によってアグリコンのc3位とc24位に糖基を付加することによって羅漢国のトリテルペンサポニンの合成が完了する[51]。モグロシドには共通のアグリコンモグロシドアルコールがあり、その違いは主にc3に結合したグルコース単位の数とc3とc24に結合したグルコース単位の数である。it—kin[50]やdai longhai[52]らの研究では、羅漢国由来のugt74ac1とugt720-269-1糖転移酵素が羅漢国アルコールのc3-ohの糖鎖化を担っていることが示されている。さらに、ugt720-269-1はモグロシドc24-ohのグリコシル化にも関与しており、ugt94-289-3はc3位とc24位でのグルコース鎖の伸長反応を担当し、最終的に5段階のグリコシル化過程を経てスイートグリコシドvを合成する(図6)。
4 .モグロシドでvivoでの分解と代謝
グリコシドの生体内での作用機序を徹底的に解析するために、近年、生体内でのモグロシドの分解と代謝に関する研究が盛んに行われている[53]。呂風来らは、ヒトの腸内細菌を使ってモグロシドを分解した結果、モグロシドiiiは、細菌の作用で、連続してc3ブドウ糖基とc24ジェンチオビオシド基を失い、ルハングクiia1とルハングクアルコールに変化することを発見した。

huang zhencong et al.[55, 56]配置Mogroside V でartificial gastric juice and intestinal bacterial liquid respectively, and tracked and analyzed its transformation products. found that the glycoside V でartificial gastric juice was hydrolyzed one によってone, and was finally converted into the aglycone; under the action of human intestinal flora, the glycoside V would undergo both deglycosylation and glycosylation reactions. The glycoside V was converted into a secondary glycoside through deglycosylation, and the glycoside V was converted into a six-glycoside by glycosylation. In vivo experiments using ネズミshowed that there are significant differences between the metabolites of mogroside V でmouse urine and feces [55]: In mouse urine, mogroside V is excreted in the form of mogroside V, while in mouse feces, mogroside V is converted to hydroxylated and isomerization products. The above research provides an important reference for clarifying the metabolism and transformation pathways of Mogroside in the body.
5議論と展望
甘味料は、食品や飲料の製造に広く使用されています。化学甘味料は無糖で低カロリーだが、消費者には受け入れられない。甘味料には化学的に合成された味があると考えている一方で、安全性や長期消費による健康への影響を懸念している。安全性の観点から、モグロシドの急性毒性はld50とgt;15 g/kg (bw)[57]であり、ames変異原性試験および非遺伝毒性試験では陰性[58]であるため、安全で毒性のない物質です。食糧の分野ではモグロシドはスクロースの自然で良い代替品です。非常に甘く、低カロリーです。消費後に体内に吸収されず、効果的にエネルギー物質の摂取量を減らすことができます[59]、これは糖尿病患者や肥満の人のニーズを満たすことができます。
薬理学的研究の面では,配糖体の低血糖活性に基づいて,科学研究者は、血糖調節における配糖体のメカニズムを研究してきました。モグロシドはインスリン分泌を刺激し、膵島細胞を修復し、糖新生を阻害し、グリコシダーゼの活性を阻害することで血糖を調節していると推測されている。このことは、グリコシドが複数の標的を持ち、多様な作用機序を持ち、体内の血糖調節に関与する複雑なシグナル経路を持っていることを示唆している。どの経路が血糖調節を主導しているのか、またどのようにして相乗的に血糖を抑制するのかについては、これまで報告されておらず、詳細な研究が待たれています。
モグロシドは潜在的な薬効があるが、高価である。モグロシドの生合成機構を明らかにし、細胞工場を用いてモグロシドを生産することは、モグロシドの大量生産の一つの可能性である。
モグロシドトリテルペン糖体の生合成においては、アセチルcoaから2,3-オキシドスクアレンへの生合成経路が高等真核生物や一部の微生物に存在する。酸化スクアレンは、ステロイドやテルペンなどの生体分子の合成の前駆体として用いられる。羅漢国では、2,3-オキシドスクアレンは一連の重要な酵素の触媒作用によって最終的にモグロシド分子に変換される。現在、生合成経路を介してモグロシドの全合成には多くの困難があり、主に3つの段階を含む:(1)スクアレンの2,3-酸化は、ククルビタジエノールを形成する;(2)ククルビタジエノールをヒドロキシル化してモマオーディノールを形成する。(3) momordinolのグリコシル化。関連する酵素遺伝子はクローニングされ、発現され、機能的に検証されていますが、これらの外来遺伝子を微生物の細胞工場に統合し、効率的かつ協調的に発現させるには、まだ多くの研究が必要です。
モグロシドの低血糖効果は、低血糖薬の開発に新たな発想を与えるもので、今後、より広範で深みのある臨床研究が求められる。モグロシドの合成はまだ初期段階にあり、多くの課題に直面しています。羅漢国のアナボリック代謝の研究と探査は、モグロシド細胞工場設立のための基礎を築くだろう。
参考:
[1]秦X, et al.Subchronic 90 口頭(に無理やり食べさせ)毒性 羅漢の研究者 郭 mogroside 抽出 in 犬か[J]。食品 2006年化学Toxicol、44:2106-2109。
村田[2] Y et al.Sweetness 特徴 of のtriterpene glycosidesで Siraitia grosvenori[J]。J Jpn Soc 食品 sci, 2006,53:527-533。
[3]金ジムよリー・ジョナサン植物化学的および薬理学的側面 siraitiagrosvenorii,羅han kuo[j]。^『仙台市史』仙台市、2012年、12:23 -239。
[4]陳DH et al.Studies and luo-hanguo(果物)からの自然な非砂糖甘味料の使用 siraitiaの grosveno - ri)か[J]。Nat Res差し込もういデヴ(天然产物研究与开发)、1992年、4
(1): 72-77。
[5] Behrens M, et al.Sweet and 旨み :収れん味自然 products, their chemosensory targets,and beyond[j]。Angew 化学Int 2011年編集、50:2220-2242。
[6]さん G .Experiment 研究 on モグロシドの肝保護効果[j]。2013年あごJ Exp Tradit救急形、19:196-200。
[7]王 Q et 増殖に対するモグロシドの影響 of 肝 stellate cell-T6 and hepatofibrosis-related 遺伝子 [J]。あご Traditi ^『仙台市史』通史編4、331- 334頁。
[8]王 Q et アル 令の規定 マウスにおけるモグロシドの免疫学的効果[j]。^ a b c d e f『仙台市史』通史編、821 - 821頁。
[9] Di R, et al.Anti-inflammatory activities of mogrosides momordicaからgrosvenori in murine マクロファージ and a マウス耳浮腫モデル[j]。J Agr食品 2011年化学、59:7474。
[10] shi d,et al。モグロシドの保護作用とメカニズム マウスにおけるlps誘発性急性肺障害について[j]。^ a b c d e f g h『仙台市史』、2014年、72 - 72頁。
[11]柳 dd,et al. siraitiagrosvenorii果実の影響 身体疲労の抽出物 in ネズミか[J]。イラン J Pharm 2013年の「Res publica、12:115-121。
[12] chen y,et al. natural food sweet- ener mogrosidesの機能的研究[j]。^中国の食品addit、2006年、1:41 -43。
鈴木。[13] 亜、et siraitia grosvenoriのトリテルペン配糖体は、ラットの腸を阻害する maltase and 鎮圧 the 上昇 in ラットにマルトースを1回経口投与した後の血糖値[j]。2005年J Agr食品化学、53:2941-2946。
[14] suzuki ya,et al siraitia grosvenori on the 自発的 糖尿病 goto-kakizakiネズミか[J]です^ a b c d e f g h i n t e r r,2007, pp . 775 -775。
[15]「姜 SYメトロ・カードとデビッド al.GAP Siraitiaの grosvenorii cul- in vitroとそのsopの確立[j]。2007年Guihaia 27: 867-872。
[16] Mo tis- sue-culturedの標準操作手順 苗 乗算 of Siraitia grosvenorii (Swingle) C.Jeffrey [J]。^『仙台市史』通史編、仙台市、2008年、209 -209頁。
〔17〕リード J et al.A 並進 synthetic biology プラットフォーム for 新規薬剤様モルのグラムスケール量への迅速なアクセス- cules[j]。2017年Metab運営者、42:185-193。
[18] Pawar RSメトロ・カードとデビッド al.Sweeteners から plants-with 強調 on stevia rebaudiana(bertoni)とsiraitia grosvenorii(swingle) [j]。^『仙台市史』通史編、仙台市、2013年、4407 -4407頁。
[19]崎XYメトロ・カードとデビッド 糖尿病性ウサギの血糖値を低下させるsiraitia grosvenori powderおよびその抽出剤の有効性試験[j]。食品Sci、2003年24:124-127。
[20] Szkudelski T .の 機構 of alloxan and streptozotocin ラット膵臓のb細胞に作用する[j]。^ a b c d e f g h i『人事興信録』、2001年、53 - 53頁。
[21] qi xy,et al. siraitia grosvenori scavからのモグロシド抽出物- in vitroでフリーラジカルを取り込み、酸化ストレスである血清を低下させる グルコース→ 脂質 levels in alloxan-induced 糖尿病 mice [J]。^『官報』第2828号、大正8年、278-284頁。
[22]陳SYメトロ・カードとデビッド morgroside interve—ning in related oxidative stress damage in pancreatic islet b パルミチン酸によって誘導される細胞[j]。^『仙台市史』通史編、仙台市、2012年、2116-2119頁。
[23]陳SYメトロ・カードとデビッドal.Progress 2型糖尿病における膵島β細胞および関連治療薬に対するoxi-dative stress damageのメカニズムで[j]。^パウサニアス、2011年10月23日、35 -35頁。
[24]張LQメトロ・カードとデビッド (c) in vitro抗酸化物質acに関する研究-シライチア・グロヴナーリ抽出物の活性。^ a b c d e f g h i 2006, pp . 227 -216。
[25] qi xy,et al. 1型糖尿病マウスの細胞免疫系に対するモグロシドを含むサイライティヤのグロブノーリエキスの影響[j]。^『仙台市史』仙台市史編纂委員会、2006年、50:32 -738。
[26]河 CWメトロ・カードとデビッド al.新鮮なマンゴーのrregulation効果-血糖値にsteenサポニン[j]。2012年Mod食品Sci技術、28:382-386。
【27】周Y et al.Insulin 分泌 モグロ-サイドの刺激効果 v and 果物 抽出物 luo 漢(漢)-漢の氏族 swingle)果物 抽出[J]。Acta pharm 2009年罪、44:1252年- 1257年。
【28】表に日焼け mj,et al.苦いメル由来のククルビタントリテルペノイドで、ampkの活性化に伴う強力な抗糖尿病作用を有する[j]。2008プランテーションMedです74:913-913。
[29] zhang ln,et al. novel small-molecule amp-activated protein kinase allosteric 促進剤 with 有益 効果 in db / dbマウス[j]。^ a b c d e f g h i、2013年8月8日。
[30] chen xb,et al. potential ampk activators of cucurbitane trit- erpenoids siraitiaからgrosvenoriiswingle[j]。Bioorgan医学 2011年化学、19:5776-5781。
[31] luo z,et al.ククルビタン型トリテルペノイドであるモグロシドvのin vitro ampk活性化効果とin vivo phar—マコキネティック からsiraitia grosvenorii 果物か[J] rsc adv,2016, 6:34 - 7041。
[32] Ag Hにする。薬理学 of α-glucosidase 抑制か[J]。Eur 投資J Clinも、1994年24:3-10だ。
[33] nalユニット~ Sメトロ・カードとデビッドal.Inhibitionのα確信によって-glucosidase抽出 一部の 強力な antidiabetic 薬用 ハーブか[J]ます学園 ^『官報』第35:29 -36。
[34]クマールはSのでらΑ植物から-glucosidase阻害薬:natu -夫か[J]糖尿病を治療できる接近。^アポロドーロス、2011年5月5日、19-28頁。
[35]夏X, et al.マウスの食後血糖値に対する、異なる成長期の果実から抽出されたフラクトゥスsiraitiae抽出物の影響[j]。2012年あごJ Exp Tradit救急形、18:166-170。
[36] song kj,et al.肝星細胞におけるモグロシドの活性化およびアポトーシスの影響[j]。あご ^『仙台市史』通史編、仙台市、2014年、48 -48頁。
〔37〕朱HL et al.Protective モグロシドexトラクトの効果 on ethanol-induced L-02 リンプ ダメージか[J]。j chin inst food sci technol,2015,15(1):13-18。
[38]さん G et al.Protective 元-ペリメンタルに対するモグロシドの効果 liver 負傷 in ネズミか[J]。あご 2018年Pharm 19: 163-165。
[39] wu y,et al 咳とエリミ- nating痰 effects of stemoninine 結合 with mogro彼らが知りたい Ⅴに ネズミか[J]。^パウサニアス、2017年7月28日、175 - 177頁。
[40]法戦馬雲メトロ・カードとデビッド al.Effect ローハン郭(siraitia grosvenoriスワングル)の 鼻 ピックアップローラ and を 行動 in ICRネズミか[J]暴れる。2005年Biol Pharmブル、28:238-241。
[41] Tiwari P et al.Plant 二次代謝結合グリコシル転移酵素(英語版):更新 on 拡大 知識 and スコープ [J]。2016年Biotechnol Adv 34:714-739。
戴[42] L et al.Exploiting the aglycon わいせつ のグリコシル-枯草菌とそのアプリケーションからのトランスフェラーゼbs-yjic 合成glycosides[J]。j biotechnol,2017,248:69-76。
[43]王 R, et al.Biotransformationのmogrosides[J]。^ a b c d e f g h『日本の歴史』学研、2018年、153-165頁。
[44]李 D et al.Cucurbitane glycosides から 生の果実 lo han kuo(siraitia grosvenori) [j]。化学Pharmブル2006年(平成18 54:1425-1428
[45] netala vr, et al.Triterpenoid saponins: a review on biosyn- thesis,applications and mechanism of their action[j]。2015年Int J Pharm Pharm Sci、7:28だ
[46]趙 CLメトロ・カードとデビッドal.Key triterpenoidの酵素により saponin biosyn-論文とその活動の誘導と遺伝子expres-植物におけるsions [j]。^『仙台市史』通史編、仙台市、2010年、5 - 7頁。
[47]孟JR et al.Cloningと ファルネシル二リン酸の配列解析 シンターゼ 遺伝子 in siraitia grosvenorii [J]。あご Tradit ハーブ 2011年薬、42:2512 - 2517。
[47]窒息Fett-Neto AGメトロ・カードとデビッド al.Biosynthesis of 植物 triterpenoid sapo- nins: genes,enzymes and their regulation[j]。Mini-Rev Org lg化学、2014年、11:292-306人である。
[49]張 J et al.Oxidation of cucurbitadienol 解媒 by siraitia gros- venoriiからのモグロシドの生合成におけるcyp87d18 [j]。plant cell physiol,2016,57:1000-1007。
[50] Itkin M, et 生合成経路 siraitia grosvenoriiの非糖、高強度甘味料mogroside v [J]。p natl acad sci usa,2016,113: e7619-e7628。
[51]吉川 S et al.Transglycosylation モグロシドvのトリットエルペン配糖体です in siraitia grosvenori、 cyclodextrin glu-カノトランスフェラーゼと甘さの質の改善 [J]。j appl glycosci,2006,52:247-252。
戴[52] L et al.Functional 特性化を推進するのは cucurbitadienol シンターゼと triterpene glycosyltransferase 関与 生で of mogrosides from siraitia grosvenorii [J]。2015植物細胞Physiolと56:1172-1182下位です。
[53] yang xr, et al。シアメノシドiの代謝物とその代謝物-ラットにおけるbutions [j]。2016年分子は、21:176。
[54] yang xw,et al ヒューマンか[J]の肠内细菌によりⅢしたものだ。J北京 univ: health sci,2007,39:657-662。
[55] lu lf,et al。モグロシドの安定性 Ⅴで 人工胃液とその代謝[j]。2015年Guihaia、6:792-795。
[56] zhou g,et al. healthおよびtype 2における天然物モグロシドvの代謝 diabetic 鼠か[J]。J ^ a b cアポロドーロス、2018年10月25日、25-33頁。
[57] nong yq,et al.モグロシドのexトラクションおよび薬理作用に関する研究の概要[j]。広西チワン族 ^ a b c d e f g hi(2008)、31頁。
[58] su xj9,et al.実験モグロシドの無毒性作用に関する研究[j]。^ food sci,2005,26(3):221-224。
59 [59] xu q,et al.経口モグロシド摂取による正常な人体血糖および肝酵素の変化に関する研究[j]。^ a b c d e f g h i、2007年3月28日、317 -317頁。




